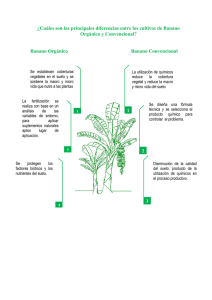
La Revista Internacional sobre Bananos y Plátanos Partenocarpia
Anuncio

La Revista Internacional sobre Bananos y Plátanos Partenocarpia en híbridos Productividad del plátano intercalado ¿ Que sería la mejor profundidad de plantación ? El sabor de los frutos de híbridos Banano orgánico en el Perú Georges Wilson Vol. 13 No.1 Junio 2004 InfoMusa Vol. 13 No.1 Foto en la portada: Pequeño vendedor en Uganda (Regis Domergue, CIRAD) INFOMUSA Vol. 13, No. 1 Editor : Red Internacional para el Mejoramiento del Banano y el Plátano (INIBAP) Jefe de Redacción: Claudine Picq Comité editorial: Anne Vézina, Jean-Vincent Escalant, Richard Markham, Nicolas Roux Con el apoyo científico de: Frédéric Bakry, Sylvio Belalcázar, Guy Blomme, Xavier Draye, Charles Staver, Jean Tchango Tchango Diagramación: Crayon & Cie Impreso en Francia ISSN: 1729-0996 Redacción: INFOMUSA, INIBAP, Parc Scientifique Agropolis II, 34397 Montpellier Cedex 5, Francia. Teléfono : + 33-(0)4 67 61 13 02; Fax: + 33-(0)4 67 61 03 34; Correo electrónico: [email protected] La subscripción es gratuita para los países en vías de desarrollo. Se agradecen contribuciones en forma de artículos y cartas al editor. La redacción se reserva el derecho de editar los artículos. INFOMUSA no se responsabiliza por el material no solicitado. Sin embargo, trataremos de responder a cada una de las peticiones. Los artículos pueden ser citados o reproducidos sin cargos, con la mención de la fuente. También se publican ediciones de INFOMUSA en francés y en inglés. Una versión electrónica esta disponible a la dirección siguiente: http://www.inibap.org/ publications/infomusa/infomusa_spa.htm Cambio de dirección: Para evitar la perdida de sus ejemplares de INFOMUSA, notifique a INIBAP con seis semanas de antelación si cambia de dirección postal. Las opiniones expresadas en los artículos son responsabilidad de sus autores y no necesariamente reflejan los puntos de vista de INIBAP. La misión de la Red Internacional para el Mejoramiento del Banano y el Plátano es aumentar de manera sostenible la productividad del banano y el plátano cultivados por pequeños productores para el consumo doméstico y mercados locales y de exportación. INIBAP es un programa del Instituto Internacional de Recursos Fitogenéticos (IPGRI), un centro Future Harvest. Contenido Crecimiento de las suspensiones celulares del cv. ‘Cau man’ Bui Trang Viet y Tran Thanh Huong 2 Efecto del Biobras-6 sobre la micropropagación del plátano FHIA-21 F.A. Jiménez Terry, D. Ramírez Aguilar y D. Agramonte Peñalver 4 Influencia de los progenitores masculino y femenino sobre la partenocarpia V. Krishnamoorthy, N. Kumar y K. Sooriyanathasundaram 7 Efectos del desmane y la distancia de siembra sobre las características productivas del plátano ‘FHIA-20’ M. Aristizábal L. 9 Efecto de la profundidad de plantación sobre la duración del ciclo de cultivo y rendimiento S.B. Bakhiet y G.A.A. Elbadri 12 Relaciones entre la capacitancia eléctrica y características de las raíces G. Blomme, I. Blanckaert, A. Tenkouano y R. Swennen 14 Productividad del plátano Falso cuerno (False horn) intercalado con el fríjol de vaca y el maíz en el sudeste de Nigeria J.O. Shiyam, B.F.D. Oko y W.B. Binang 18 Efecto del deshije sobre la resistencia de FHIA-23 y SH-3436-9 a enfermedades y plagas A. Vargas y M. Guzmán 20 Evaluación de nuevos híbridos de banano contra la enfermedad de la Sigatoka negra V. Krishnamoorthy, N. Kumar, K. Angappan y K. Soorianathasundaram 25 Cualidades organolépticas de los frutos de los híbridos SH-3640 y CRBP-39 S. Coulibaly y C. Djédji 27 Focus sobre los bananos silvestres 31 Focus sobre banano orgánico 32 In memoria de Georges Wilson 34 Tesis 35 Noticias de Musa 38 Forum 44 Subiendo los estándares científicos Editorial D e vez en cuando nos plantean considerar la idea de hacer de INFOMUSA una revista académica arbitrada. En los últimos tiempos, este planteamiento surgía más a menudo ya que el avance de la carrera de un científico está ligado a la cantidad de artículos publicados en las revistas académicas reconocidas. Nuestra posición siempre ha sido la de resistir a este cambio. Sentimos que el papel de INFOMUSA es el de informar a los miembros de la comunidad de investigación y desarrollo de lo que está pasando en el mundo de los bananos, aún si la investigación de la que se informa no es necesariamente ‘vanguardista’ desde el punto de vista científico o no rindió los resultados esperados. Sin una publicación como INFOMUSA, creemos que los investigadores bananeros perderían mucha información valiosa que permanecería enterrada en la llamada literatura gris. Los bananos se cultivan en diversos ambientes y el sentido de déjà vu es inevitable, igual que muchas variaciones sobre el mismo tema, como por ejemplo el cribado de germoplasma para la resistencia a las plagas y enfermedades comunes, las cuales se desarrollan por diferentes investigadores en sus propias situaciones. Los resultados negativos son las especies aún más olvidadas y raramente encuentran su salida en las revistas académicas respetadas. Sin embargo, los resultados negativos son extremadamente útiles para descartar las hipótesis erróneas. Informando sobre ellos prevenimos la duplicación de los esfuerzos innecesarios y la pérdida de recursos que en los países en vías de desarrollo, donde se realiza la mayor parte de las investigaciones de Musa, son particularmente escasos. Aunque queremos mantener a INFOMUSA amigable con los autores, también sentimos que el hecho de brindar la información a nuestros lectores no debe ser a expensas de la integridad científica. La información es de poco uso si los lectores no entienden de donde proviene o si no se realizaron análisis estadísticos de los datos. No obstante las reglas de la investigación científica y su presentación, de las cuales depende nuestra confianza de utilizar los resultados publicados, no son obvios intuitivamente. La mayoría de los autores, aún los que hablan en uno de los tres idiomas en las cuales se publica la revista, necesitan ayuda para presentar su trabajo, especialmente los relacionados al análisis de los datos. Tratamos de asegurar que los artículos que publicamos cumplan con los requisitos básicos de informes científicos, pasando los manuscritos a los miembros de nuestro comité editorial para una revisión informal, en base de lo cual los textos a menudo se regresan a los autores para que los mejoren. La cuestión con la cual nos enfrentamos ahora es cuan lejos podemos ir para mejorar la calidad de INFOMUSA sin sacrificar lo que ustedes, los lectores, aprecian en ella. Anteriormente hemos discutido en contra una INFOMUSA académica formalmente arbitrada, ¿pero, puede ser que ustedes sienten de manera diferente? Por favor, hágannos saber que piensan ustedes al contestar el cuestionario adjunto, el cual ha sido diseñado para descubrir su opinión sobre los cambios que hemos hecho recientemente a INFOMUSA y qué podemos hacer para producir una revista que cumpla mejor con sus necesidades. También pueden escribirnos. En este número, estamos inaugurando la sección Forum, que está abierta a todos nuestros lectores. Esperamos que ustedes la utilicen para comentar sobre los artículos publicados o inicien debates sobre los tópicos relevantes. El sitio de Internet de INIBAP también esta cambiando. Fue expandido para incluir una sección sobre los usos de los bananos y sus productos, información general sobre los bananos dirigida al público en general y un sitio sobre genómica para informar sobre el progreso hecho en la decodificación de los genomas del banano y de Mycosphaerella. Los editores InfoMusa - Vol. 13 - No.1 1 Suspensiones celulares Crecimiento de las suspensiones celulares del cv. ‘Cau man’ Bui Trang Viet y Tran Thanh Huong L a multiplicación de los bananos in vitro se realiza principalmente a través de la proliferación de los meristemas vegetativos. El desarrollo reciente de suspensiones de células embriogénicas abre la posibilidad para la producción masiva de plantas de banano a bajo costo (Haicour et al. 1998). Varios estudios sobre las suspensiones celulares se han realizado en la Universidad Nacional de Vietnam en la ciudad de Ho Chi Minh (Bui Trang Viet et al. 2000, Tran Thanh Huong y Bui Trang Viet 2000, Cung Hoang Phi Phuong y Bui Trang Viet 2000, Tran Thanh Huong y Bui Trang Viet 2003). En este trabajo informamos sobre el crecimiento de una suspensión celular a partir de las flores masculinas inmaduras de un cultivar cultivado ampliamente en Vietnam, el ‘Cau man’. Materiales y métodos Se recolectaron flores masculinas inmaduras y luego se colocaron en el medio MA1 (Escalant et al. 1994, Shii et al. 1992). La suspensión celular fue iniciada colocando el callo de 4 meses de edad en el medio MA2 líquido (Escalant et al. 1994, Shii et al. 1992) complementado con 35 g/L de sacarosa, 1 mg/L de 2,4-D, y 15 mg/L de ácido ascórbico. Los cultivos se mantuvieron en frascos de Erlenmeyer de 100 ml que fueron colocados en un temblador giratorio a 80 rpm y cultivados a 28°C bajo 1000 lux (fotoperíodo de 12 h). El medio viejo de cultivo fue reemplazado con un medio fresco cada dos semanas. Después de dos meses, la suspensión contenía una mezcla de manojos de células y células individuales. Después de dos semanas de subcultivo, la suspensión celular fue filtrada a través de Figura 1. Tasa de respiración durante el crecimiento de la suspension celular. Resultados y discusión 800 600 Respiración (�l O2/g/h) filtros de metal de 0.8 mm. El crecimiento de las células fue evaluado midiendo el volumen de células establecidas (SCV) después de 5 minutos de sedimentación (Gómez et al. 2000, Schoofs 1997). Durante el crecimiento de las suspensiones celulares se midió la tasa de respiración. También se midieron cada semana los cambios en el pH y las concentraciones de oxígeno y las sales totales en el medio de crecimiento. El medio de crecimiento fue separado de la suspensión celular rotando a 1000 rpm durante 5 minutos. La presencia del ácido indoleacético (AIA), una auxina, y del ácido abscísico (ABA) se detectó utilizando cromatografía en capa fina en gel de sílice (60 F254, 105554, Merk). Los cromatogramas fueron revelados en cloroformo:metanol:ácido acético (80:15:5 v/v). Las sustancias reguladoras del crecimiento fueron visualizadas bajo la luz UV, de acuerdo a Yokota et al. 1980. La identificación se realizó comparando con los estándares de referencia conocidos de AIA y zeatina. Para medir el nivel de AIA se utilizó la prueba de crecimiento recto coléoptilos Oryza. Esta prueba está basada en la habilidad de la auxina de estimular la elongación de las secciones de coléoptilos que flotan en el medio líquido. La actividad de zeatina fue detectada utilizando una prueba basada en la expansión dependiente de la citoquinina de los cotiledones de pepino (Meidner 1984). Alícuotas de 5 ml de suspensión celular fueron filtradas a través de un tamiz de 800 µm y inoculadas en 5 ml del medio MA3 sólido complementado con 0.2 mg/L de NAA, 0.1 mg/L de quinetina y 0.05 mg/L de zeatina (Escalant et al. 1994, Shii et al. 1992). 400 200 0 0 7 14 Días 2 21 28 El crecimiento de la suspensión celular en el tiempo siguió una curva sigmoidea. Un período de crecimiento lento, hasta el séptimo día, fue seguido por un período de crecimiento rápido entre el 7o y 21r días. Este crecimiento rápido fue seguido por una fase estacionaria. La respiración aumentó durante el período de crecimiento rápido y disminuyó antes de la fase estacionaria (Figura 1). La influencia de la densidad de inoculación inicial sobre el crecimiento de la suspensión celular se estudió examinando tres volúmenes de células establecidas: 50, 75 y 100 µl/ml del medio. El mejor porcentaje de células embriogénicas con un citoplasma InfoMusa - Vol. 13 - No.1 embriogénica de las células del ‘Cau man’ pero las condiciones actuales de cultivo deben ser mejoradas con el fin de obtener embriones somáticos bien formados. Figura 2. Manojos de células después de dos semanas de cultivo. Agradecimiento Este estudio recibió apoyo de una subvención del Consejo Nacional de Viet Nam sobre los problemas fundamentales en ciencias naturales. 6 6 5 5 4 4 Oxigeno Sales totales pH 3 2 3 2 1 1 0 0 0 7 pH Figure 3. Cambios en el pH y concentraciones de las sales totales y oxigeno en el medio de cultivo. Concentración (mg/L) denso y núcleo grande (Figura 2) se observó a 75 µl/ml. Después de reemplazar el viejo medio de cultivo con un medio fresco, la suspensión resultó ser heterogénea y contenía una mezcla de manojos celulares y células individuales. Los manojos de células se dividieron en dos grupos basándose en su diámetro: pequeño (menor de 200 µm), medio (200-800 µm) y grande (mayor de 800 µm). El crecimiento de la suspensión celular fue evaluado midiendo el número de estas células y manojos. El número máximo de manojos pequeños y medianos (48 000±147 y 1820±120, respectivamente) se observaron al 14° día, mientras que el número máximo de células individuales (77 280±2219) y manojos grandes de células (338±18) se observaron al 21r día. Durante la fase de crecimiento rápido, un gran número de pequeños manojos se formó de los manojos grandes. El SCV total fue muy bajo cuando el cultivo se establecía a partir de manojos pequeños (Tabla 1). El pH y las concentraciones de oxígeno y sales totales en el medio de crecimiento disminuyeron durante el crecimiento de la suspensión celular (Figura 3). Utilizando cromatografía monodimensional en capa fina, la actividad de las auxinas correspondiente al AIA se identificó a Rf 0.84-0.89, y la actividad de la citoquinina correspondiente a la zeatina se observó a Rf 0.67-0.74. Los niveles de estas hormonas de crecimiento durante el crecimiento de la 14 Días 21 28 Tabla 1. Volumen de células establecidas (µl/ml del medio) de las suspensiones celulares después de 14 días de cultivo. Tamaño final de los manojos de célula 200-800 µm Más de 800 µm 27.4 ±2.4 3.9 ±3.2 20.2 ±2.0 58.1 ± 5.9 42.9 ± 1.7 50.4 ±1.2 25.4 ±2.8 67.2 ± 5.0 suspensión celular se muestran en la Figura 4. Los resultados muestran que el crecimiento de la suspensión celular aumentó con una concentración aumentada de zeatina pero los altos niveles de AIA inhibieron este crecimiento. Con 0.2 mg/L de ácido naftaleneacético (ANA), 0.1 mg/L de quinetina y 0.05 mg/L de zeatina, los embriones somáticos se desarrollaron normalmente hasta llegar a la etapa globular (después de 8 días de cultivo, lo más temprano) pero su desarrollo subsiguiente fue suprimido. La ausencia de la biopolaridad de los embriones fue causada por una pobre individualización de los brotes y meristemas radicales. Se encontraba presente una epidermis pero no hubo una organización interna clara. Los resultados proporcionan evidencia de la capacidad InfoMusa - Vol. 13 - No.1 Total 54.0 ± 5.1 97.9 ± 8.6 100.8 ±10.0 150.1 ±12.0 Figura 4. Cambios en el pH y concentraciones de la sales totales y oxígeno en el medio de culivo. 160 9 AIA 8 Zeatina 7 120 Crecimiento de la suspensión celular 6 5 80 4 3 40 2 1 0 0 0 7 14 Días 21 3 Volúmen de células establecidas (�/ml del medio) Menos de 200 µm 22.7 ±1.4 19.6 ±2.0 7.5 ± 0.6 57.4 ± 4.3 Concentraciones de las hormonas de crecimiento (�g/l) Tamaño inicial de los manojos de celulas Menos de 200 µm 200–800 µm Menos de 800 µm Más de 800 µm 28 Referencias Bui Trang Viet trabaja en el Department of Plant Physiology, University of Natural Sciences, Vietnam, e-mail: [email protected] y Tran Thanh Huong en la Universidad Nacional, 227 de la calle Nguyen Van Cu, distrito 5, Ciudad de HoChiMinh, Vietnam, e-mail: [email protected] Cultivo de tejidos Bui Trang Viet, Cung Hoang Phi Phuong, Tran Thanh Huong & Pham Thanh Luong. 2000. Callus and cell suspension initiation from male flowers of M. balbisiana cv. Hot and M. cavendishii cv. Gia cui (abstract in English). Viet Nam National University-HCM City, College of Natural Sciences, May-2000. Proceedings of Symposium, pp. 114-118. Cung Hoang Phi Phuong & Bui Trang Viet. 2000. Development of embryos and callus initiation from immature embryos of Musa balbisiana (abstract in English). J. Science & Technology Development 3(56):38-44. Escalant J.V., C. Teisson & F. Cote. 1994. Amplified somatic embryogenesis from male flowers of triploid banana and plantain cultivars (Musa spp.). In Vitro Plant Cell and Dev. Biol. 30:181-186. Gómez R.K., T. Gilliard, L.A. Barranco & M. Reyes. 2000. Embryogenesis somática en medios líquidos. Maduración y aumento de la germinación en el cultivar híbrido FHIA-18 (AAAB). INFOMUSA 9(1):12-16. Haicour R., V. Bui Trang, D. Dhed’a , A. Assani, F. Bakry & F.X. Cote. 1998. Banana improvement through biotechnology-ensuring food security in the 21st century (Abstract in English). Cahiers Agricultures 7:468-475. Meidner H. 1984. Class Experiments in Plant Physiology. George Allen and Unwin (London). 169pp. Schoofs H. 1997. The origin of embryogenic cells in Musa. Dissertationes de Agricultura. Catholic University of Leuven, Belgium. 258p. Shii C.T., S.S. Ma, I.C. Huang & W.H. Ching. 1992. Somatic embryogenesis and plantlet regeneration in suspension cell cultures of triploid banana (Musa AAA, subgroup Cavendish). Pp. 21-22 in Abstracts of International Symposium on Recent Development in Banana Cultivation Technology. Fifth meeting of International Group on Horticultural Physiology of Banana, Pingtung Taiwan, 14-18 Dec. Tran Thanh Huong & Bui Trang Viet. 2000. Effects of various antioxidants on Musa cavendishii cell suspension culture (abstract in English). Viet Nam National University-HCM City, College of Natural Sciences, May-2000. Proceedings of Symposium, pp.17-23. Tran Thanh Huong & Bui Trang Viet. 2003. Growth of the cell suspension of Musa paradisiaca L. cv. Cau Man (abstract in English). Viet Nam National UniversityHCM City, College of Natural Sciences. Proceedings of Symposium on Natural Sciences, October 2002, pp. 423-430. Yokota T., N. Murofushi & N. Takahashi. 1980. Molecular aspects of plant hormones. Pp. 113-201 in Encyclopedia of plant physiology, 9 (J. MacMillan, ed.). Springer New York Efecto del Biobras-6 sobre la micropropagación del plátano FHIA-21 F.A. Jiménez Terry, D. Ramírez Aguilar y D. Agramonte Peñalver L a baja tasa de multiplicación, ocurrencia de alta oxidación de los explantes y crecimiento lento son algunos problemas que encontrados en la micropropagación de los plátanos y en particular del híbrido FHIA-21 (AAAB). El Biobras-6, un análogo de brasinoesteroide pudiera sustituir a algunos reguladores del crecimiento empleados en el cultivo de tejidos (Gómez et al. 2000). Se realizó el presente trabajo con el objetivo de evaluar el efecto de Biobras-6 sobre la micropropagación in vitro de FHIA-21. Materiales y métodos Los ensayos se realizaron en el Instituto de Biotecnología de las Plantas (IBP) con plantas in vitro procedentes de tres subcultivos del clon de plátano híbrido FHIA-21. El instrumental utilizado se esterilizó en un horno de calor seco a 180ºC y los medios de cultivo en autoclaves a una temperatura de 121ºC y 1.2 kg/cm² de presión durante 20 minutos. El material vegetal se colocó en cámaras de 4 luz natural con valor promedio de 4500 lux y temperatura de 28±2ºC. Fase de multiplicación El medio basal de multiplicación fue el compuesto por las sales propuestas por Murashige y Skoog (sales MS), 1.0 mg/L de tiamina, 3% sacarosa y 8% de agar. Cada frasco contenía 30 ml del medio de cultivo y 10 explantes. Se realizaron 10 réplicas por tratamiento. En las variantes de medios líquidos se redujeron las sales MS al 70% y se aplicó 15 ml de medio por frasco de 250 ml de capacidad. Se realizaron tres subcultivos cada 21 días. Los brotes axilares fueron separados y los tejidos con oxidación fenólica extraídos y el cormo de las plantas completamente formadas cortados transversalmente. . Se realizaron dos experimentos para los estados físicos semisólido y líquido en la fase de multiplicación. Se evaluaron diferentes combinaciones de dos concentraciones de citokinina 6-BAP (2.0 y 4.0 mg/L) y de Biobras-6 (0.01 y 0.05 mg/L) (Tabla 1). La InfoMusa - Vol. 13 - No.1 tasa de multiplicación, el número de brotes por explante, el número de raíces por explante y la longitud de las plantas fueron registrados después de 21 días. Fase de enraizamiento En la fase de enraizamiento se incrementó de 4% la concentración de sacarosa el medio basal. Las diferentes concentraciones de Biobras-6 evaluadas en la fase de enraizamiento se presentan en la Tabla 2. La tasa de multiplicación, el número de brotes por explante, el número de raíces por explante y la longitud de las plantas fueron registrados a los 28 días. Fase de aclimatización En la fase de aclimatización, se aplicó la inmersión de las raíces de las plantas in vitro en varias soluciones de Biobras-6 y el acido naftalenacetico (ANA), durante 10 minutos antes de la plantación (Tabla 3) Las vitroplantas se colocaron en cajas de polieturano de 70 alvéolos y dimensiones de 69 cm x 45 cm. A los 45 días del ciclo de aclimatización, los datos registrados fueron el número de hojas y raíces por explante, el peso fresco y el peso seco de la planta. Todos los datos de los experimentos fueron procesados a través del paquete estadístico Statistical Program Scientific System (SPSS) versión 9.0 para Windows. Con el mismo se efectuó la previa comparación de los supuestos de homogeneidad de varianzas y normalidad de los datos, y posteriormente los análisis multifactorial de varianza con prueba de rango múltiple de Duncan. Resultados y discusión Fase de multiplicación Como se puede observar en la Tabla 4, el número de brotes por explante y tasa de multiplicación en el medio semisólido fueron significativamente mas altos en los tratamientos 5 y 6 que contenían 4 mg/L 6-BAP + 0.01 mg/L y 0.05 mg/L Biobras-6 respectivamente. Sin embargo, el número de brotes por explante en el testigo, que contenía 4 mg/L 6-BAP pero no Tabla 1. Concentraciones de biobras-6, 6-BAP y ácido indoleacético (AIA) en la fase de multiplicación. Tratamientos (mg/L) 1 2 3 4 5 6 Testigo 6-BAP (mg/L) 0 0 2 2 4 4 4 Biobras-6 (mg/L) 0.01 0.05 0.01 0.05 0.01 0.05 - Tabla 2. Concentraciones de biobras-6, 6-BAP y ácido indoleacético (AIA) en la fase de enraizamiento. Tratamientos 1 2 3 4 5 6 Testigo Biobras-6 (mg/L) 0.01 0.05 0.01 0.05 0.01 0.05 - AIA (mg/L) 0.65 0.65 1.3 1.3 1.3 Tabla 3. Concentraciones de biobras-6, 6-BAP y ácido naftalenacético (ANA) en la fase de aclimatización. Tratamientos 1 2 3 4 Testigo Biobras-6 (mg/L) 0.01 0.01 0.05 0.05 ANA (mg/L) 10 10 10 Biobras, no fue significativamente diferente del valor obtenido en los tratamientos 5 y 6. La mayor longitud del tallo de las plantas se observó en los tratamientos 1 y 2 con Biobras-6 pero sin la citoquinina 6-BAP lo que sugiere un efecto auxínico. Los explantes de estos tratamientos también presentaron los valores los más bajos en cuanto a la tasa de multiplicación y los mas altos en cuanto al número de raíces. Los resultados obtenidos sugieren que el Biobras-6 se puede utilizar como sustituto del AIA en el medio de cultivo de multiplicación del plátano FHIA-21. Los resultados con el medio de cultivo liquido Tabla 4. Efecto en FHIA-21 de varias concentraciones de reguladores de crecimiento en un medio semisólido para la fase de multiplicación (ver Tabla 1 para los detalles de los tratamientos). Tratamientos 1 2 3 4 5 6 Testigo Númer de brotes por explante Tasa de multiplicación Número de raíces por explante Longitud del tallo de las plantas (cm) 1.3 c 1.7 c 2.6 b 2.9 b 3.4 a 3.8 a 3.1 a 1.1 d 1.1 d 2.2 c 2.5 c 3.1 a 3.5 a 2.8 b 6.1 c 7.9 c 0.6 a 1.3 b 0a 0a 0.2 a 2.9 ab 3.2 a 2.5 bc 2.7 b 2.3 c 2.6 b 2.5 bc Letras diferentes en un indicador difieren estadísticamente (p< 0.05). InfoMusa - Vol. 13 - No.1 5 AIA (mg/L) 0.65 (Tabla 5) siguen el mismo patrón que con el medio semisólido. Se observaron efectos de los brasinoesteroides en la elongación, la división celular, el desarrollo vascular y reproductivo (Núñez 2000). La utilización del Biobras-6 durante el cultivo in vitro del banano FHIA-18 tuvo un efecto en la germinación de embriones somáticos (Gómez et al. 2000). Un medio de cultivo conteniendo ambos Biobras-6 y 6-BAP es más favorable a la división celular y la formación de brotes y permite incrementar la tasa de multiplicación sin el aspecto negativo de la formación de raíces observado cuanto el Biobras-6 esta utilizado solo. Fase de enraizamiento Los mejores valores para el numero de raíces por explante, longitud del tallo, peso fresco y peso seco de la planta se observaron en los tratamientos combinando el Biobras-6 y el AIA, y los valores de longitud del tallo y peso fresco de la planta no fueron significativamente diferentes de aquellas obtenidas con el AIA solo (Tabla 6). Resultados anteriores relativos al proceso de micropropagación masiva de plátanos y bananos demostraron que el Biobras-6 es capaz de sustituir al 6-BAP en la fase de establecimiento (Barranco 2001) y al AIA en la fase de enraizamiento (Nuñez 2000). Fase de aclimatización Los resultados nos muestran que las variables altura de las plantas y peso fresco de las plantas en la fase de aclimatización son superiores en el tratamientos 4 (0.05 mg/L Biobras-6 + 10 mg/L ANA). Tabla 5. Efecto en FHIA-21 de varias concentraciones de reguladores de crecimiento en un medio líquido para la fase de multilicación (Ver Tabla 1 para los detalles de los tratamientos). Tratamientos 1 2 3 4 5 6 Testigo Número de brotes por explante Tasa de multiplicación Número de raíces por explante Longitud del tallo de las plantas (cm) 1.1 c 1.1 c 2.2 b 2.4 b 2.7 a 3.0 a 2.7 a 1.0 d 1.0 d 1.8 c 2.1 c 2.5 a 2.8 a 2.5 a 5.3 c 5.5 c 0.3 a 1.3 b 0a 0a 0.2 a 3.1 a 3.2 a 2.6 b 2.7 b 2.4 b 2.7 b 2.5 b Letras diferentes en un indicador difieren estadísticamente (p< 0.05). Tabla 6. Efecto en FHIA-21 de varias concentraciones de reguladores de crecimiento en un medio para la fase de enraízamiento (Ver Tabla 2 para los detalles de los tratamientos). Tratamientos Número de hojas por explante 1 3.5 2 3.7 3 3.5 4 3.8 5 3.7 6 3.9 Testigo 3.8 Número de raíces por explante 4.9 c 5.1 c 5.6 b 5.8 ab 6.3 a 6.2 a 5.6 b Longitud del tallo (cm) 3.8 b 4.0 b 4.2 b 4.7 a 4.9 a 4.8 a 4.8 a Peso fresco de la planta (g) 1.8 b 2.0 b 2.4 a 2.6 a 2.6 a 2.4 a 2.5 a Peso seco de la planta (g) 0.30 d 0.36 c 0.39 bc 0.41 ab 0.43 a 0.46 a 0.39 bc Letras diferentes en un indicador difieren estadísticamente (p< 0.05). Referencias Tabla 7. Efecto en FHIA-21 de varias concentraciones de reguladores de crecimiento en la fase de aclimatización. (VerTabla 3 para los detalles de los tratamientos). Tratamientos Los autores trabajan en el Instituto de Biotecnología de las Plantas (IBP), Universidad Central de Las Villas, Carretera a Camajuani km 51⁄2, Santa Clara, Villa Clara, CP 54830, Cuba. 1 2 3 4 Testigo Longitud del tallo (cm) 12.3 d 12.8 b 12.5 c 13.2 a 12.6 c Peso fresco (g) 109.9 d 117.6 b 112.4 c 119.0 a 111.2 cd Letras diferentes en un indicador difieren estadísticamente (p<0.05). 6 Barranco L.A. 2001. Desarrollo de la embriogénesis somática en medios líquidos (Musa AAA cv. Gran enano). Taller Internacional de Biotecnología Vegetal. Libro de resúmenes. Centro de Bioplantas. Ciego de Avila. Cuba. . Gómez R., T. Gillard, L.A. Barranco & M. Reyes. 2000. Embriogénesis somática en medios líquidos. Maduración y aumento de la germinación en el cultivar híbrido FHIA-18. INFOMUSA 9(1):12-16. Murashige T & F Skoog.1962. A revised medium for rapid growth and biossays with tobacco tissue culture. Plant Physiology 15:473-479. Núñez M. 2000. Tendencias actuales del uso de los brasinoesteroides en la agricultura. P. 24 in XII Seminario Científico INCA. Instituto Nacional de Ciencias Agrícolas, La Habana, Cuba. InfoMusa - Vol. 13 - No.1 Influencia de los progenitores masculino y femenino sobre la partenocarpia Mejoramiento V. Krishnamoorthy, N. Kumar y K. Sooriyanathasundaram E l hecho de que los cultivares triploides no tienen semillas los hace comestibles pero también representa una limitación cuando se trata de mejorar su rendimiento y resistencia a los estrés bióticos. Los mejoradores siempre tratan de obtener híbridos partenocárpicos con resistencia mejorada. Simmonds (1952, 1953 y 1959) reportó que la partenocarpia es controlada por tres genes dominantes complementarios. El presente estudio se realizó para investigar la germinación de las semillas y la partenocarpia en la progenie resultante de los cruzamientos entre los diploides, triploides y diploides con triploides. Materiales y métodos El trabajo se llevó a cabo en el Colegio Hortícola e Instituto de Investigaciones, Universidad Agropecuaria de Tamil Nadu en Coimbatore. Los triploides y diploides utilizados en el estudio se presentan en la Tabla 1, y los híbridos diploides sintéticos en la Tabla 2. Con excepción de ‘Rasthali’, ‘Nendran’ y ‘Ney poovan’, que son cultivares masculinos estériles, todos los otros cultivares fueron utilizados como progenitores femeninos y masculinos. ‘Robusta’ fue utilizado como progenitor masculino. Tabla 1. Cultivares utilizados en los cruzamientos. Cultivares triploides Nendran Red banana Robusta Rasthali Bareli chinia Karpooravalli Cultivares diploides Ambalakadali Anaikomban Eraichivazhai Matti Namarai Nivediyakadali Pisang lilin Sannachengadali Tongat Ney poovan Grupo Subgrupo AAA AAA AAA AAB ABB ABB Plátano French Red Cavendish gigante Silk Pisang awak AA AA AA AA AA AA AA AA AA AB Tabla 2. Parentesco de los híbridos diploides sintéticos utilizados en los cruzamientos. H-110 H-201 Grupo AA AB Progenitores Matti × Tongat (Bareli chinia × Pisang lilin) × Robusta InfoMusa - Vol. 13 - No.1 Se realizó un total de 7830 cruzamientos, representando 71 combinaciones. Los cruzamientos se realizaron entre las 7.30 am y 10.30 am. Las anteras sin abrirse de los progenitores masculinos fueron recolectadas de las flores abiertas el día de cruzamiento. Las anteras fueron retorcidas y forzadas a abrirse. El polen se recolectó y se untó sobre el estigma de las flores femeninas utilizando un cepillo No. 2. El estigma receptor era pegajoso al tacto, mientras que el estigma que perdió la receptividad era de color azuloso o marrón. Las flores fueron cubiertas con una bolsa de papel perforada y marcadas con la información sobre la fecha del cruzamiento, la combinación parental y el número de dedos cruzados. Las bolsas fueron removidas cinco días más tarde. Los racimos fueron cosechados después de que uno o dos dedos maduraron in situ. Las semillas se extrajeron manualmente de la pulpa y se clasificaron como ‘buenas’ o ‘malas’ después de una inmersión en agua. Las semillas flotantes se consideraron malas y se descartaron. Las semillas fueron remojadas en agua por ocho días y sembradas en una mezcla de tierra esterilizada (Rowe y Richardson 1975). Ellas se mantuvieron en una cámara de neblina y su germinación fue vigilada de cerca. Sólo 312 plantones híbridos fueron obtenidos de 18 de las 71 combinaciones intentadas. Los híbridos fueron sembrados en el campo y al momento de la floración las flores femeninas fueron cubiertas con una bolsa de papel para prevenir la polinización natural. Un mes después de la floración, se observó el desarrollo de la pulpa en una sección transversal en las flores femeninas. La ploidia de los híbridos fue evaluada en dos momentos diferentes. La densidad de los estomas y el tamaño y la cantidad de cloroplastos por un par de células de guarda fueron medidas en la etapa de plántulas. Los híbridos que tenían de 62.6 a 109.6, de 50.4 a 56.9 y de 19.7 a 35.1 estomas por milímetro cuadrado fueron clasificados respectivamente como diploides, triploides y tetraploides (Sathiamoorthy 1973). Se contó el número de cloroplastos en el par de células de guarda utilizando el método sugerido por Tenkouano et al. (1998). Las plantas con una densidad de cloroplastos entre 10.1 y 10.3, 13.1 y 13.3 y entre 15.5 y 15.8 fueron clasificadas respectivamente como diploides, triploides y tetraploides. 7 Frutas del híbrido tetraploide resultando de un cruzamiento entre ‘Karpooravalli’ y ‘Red banana.’ Posteriormente, se contó el número de cromosomas en las puntas de las raíces. El número de cromosomas se determinó utilizando una versión ligeramente modificada del método descrito en Dolezel et al. (1997). La composición genómica de los nuevos híbridos fue evaluada utilizando un método de anotación morfológica (Simmonds y Shepherd 1955). Resultados y discusión De las 71 combinaciones intentadas, sólo 23 produjeron semillas para un total de 1096 (Tabla 3). De estas, 1003 fueron viables, o no flotantes, y las restantes 93 fueron semillas flotantes y se consideraron semillas malas. El número promedio de semillas por fruta fue 1.23. Las tasas de germinación más altas fueron observadas con el H-201 polinizado abiertamente y los cruzamientos H-201 × H-110 y ‘Karpooravalli’ × ‘Red banana’. Ninguna de las semillas obtenidas de los cruzamientos entre ‘Anaikomban’ y ‘Pisang lilin’, ‘Anaikomban’ y ‘Eraichivazhai’, ‘Ambalakadali’ y ‘H-110’, ‘Karpooravalli’ y ‘Anaikomban’ y entre ‘Red banana’ y ‘Pisang lilin’ germinó. De las 1003 semillas viables, 312 semillas que representaban 18 combinaciones, germinaron. El tiempo tomado para la germinación varió de 15 días (polinización abierta de H-201) a 54 días (‘Anaikomban’ × H-110). Se observaron un pobre establecimiento de semillas y una alta ocurrencia de semillas vacías o parcialmente llenas. Simmonds (1952, 1959) sugirió que la ocurrencia de las semillas parcialmente llenas, germinación fracasada y la mortandad prematura de las plántulas podría atribuirse a los eventos genéticos y citológicos que tienen lugar inmediatamente después de la fertilización del banano. La tasa de germinación relativamente buena podría deberse al hecho de que esta tuvo lugar en los meses de febrero y marzo, durante los cuales prevalecen días soleados y noches frescas. La experiencia de este departamento dice que se debe evitar los meses más fríos (de noviembre a enero) y los meses más calurosos (mayo y junio). Sathiamoorthy (1987) reportó que la germinación fue muy baja y errática en invierno. La ploidia fue estimada mediante la apariencia fenotípica y confirmada por la densidad de estomas o por el número de cloroplastos por el par de células de guarda y conteo de cromosomas en las puntas de las raíces. En el presente estudio, la densidad de estomas no concurrió con los niveles de ploidia en informes anteriores (Sathiamoorthy 1987) así que, para la evaluación de la ploidia se utilizaron las Tabla 3. Número y caracterización de los híbridos obtenidos de las 23 combinaciones exitosas. Combinación Número de dedos cruzados AA × AA Anaikomban × Pisang lilin Anaikomban × Eraichivazahi Anaikomban × H-110 Ambalakadali × Anaikomban Ambalakadali × H-110 Matti × Pisang lilin AB × AA H-201 × Eraichivazhai H-201 × Anaikomban H-201 × Ambalakadali H-201 × Pisang lilin H-201 × Nivediyakadali H-201 × H-110 H-201 (polinizados abiertamente) Triploide × Diploide Karpooravalli × Pisang lilin Karpooravalli × H-110 Karpooravalli × Nivediyakadali Karpooravalli × Eraichivazhai Karpooravalli × Ambalakadali Karpooravalli × Anaikomban Red Banana × Pisang lilin Triploide × Triploide Karpooravalli × Red banana Karpooravalli × Robusta Karpooravalli (polinizados abiertamente) Días hasta la germinación Semillas Número de híbridos obtenidos Hundidas Flotantes 31 30 33 15 30 30 2 1 7 1 2 30 64 102 30 79 36 53 64 P* Ploidia 2x 3x 4x 5x 54 51 1 1 1 1 1 26 4 4 4 142 105 5 340 2 73 50 31 7 1 11 2 4 5 29 18 18 18 26 18 15 49 15 1 115 1 33 37 38 12 1 90 1 17 1105 630 135 107 155 80 137 25 9 10 16 54 1 0 3 32 34 37 35 34 7 1 3 1 1 233 150 195 87 80 33 6 13 11 26 28 26 33 8 2 3 2 1 5 3 1 NP** 1 6 1 2 25 1 16 4 6 1 3 1 1 3 48 13 1 114 1 29 4 1 3 1 4 1 29 5 4 3 16 1 17 7 * Numero de híbridos partenocárpicos ** Numero de híbridos no partenocárpicos 8 InfoMusa - Vol. 13 - No.1 variaciones de las densidades de los estomas de los progenitores. Basándose en este criterio, 299 híbridos (197 diploides, 1 triploide y 101 tetraploides) se clasificaron en el rango de los cultivares parentales, mientras que 15 resultaron fuera. Al utilizar el método del número de cloroplastos, 15 híbridos no se clasificaron ni como diploides, ni triploides ni tetraploides. Los conteos de los cromosomas en las puntas de las raíces se realizaron en 48 híbridos y los 15 híbridos que no pudieron ser clasificados ni como diploides, ni triploides ni tetraploides. El nivel de la ploidia de 33 híbridos concordó con los resultados de la densidad de estomas y 13 de los 15 híbridos tuvieron 55 cromosomas. De los 312 híbridos obtenidos, 36 fueron partenocárpicos. De los 6 híbridos obtenidos de los cruzamientos AA × AA, 5 fueron partenocárpicos (Tabla 3). La hibridación de un AB con un AA rindió 8 híbridos partenocárpicos de los 214 híbridos. Los cruzamientos triploide × diploide dieron 13 híbridos, de los cuales solo 4 fueron partenocárpicos, mientras que los cruzamientos triploide × triploide produjeron 41 híbridos, de los cuales 17 fueron partenocárpicos. En el caso de los cruzamientos entre el H-201 con los diploides AA, muchos híbridos demostraron ser no partenocárpicos, sugiriendo la influencia del progenitor femenino altamente fértil. Viendo el pedigrí del híbrido H-201, parece que ‘Bareli chinia’, que fue utilizado como progenitor femenino, podría explicar la no partenocarpia observada en este estudio. Los híbridos partenocárpicos obtenidos en los cruzamientos entre el H-201 y un progenitor diploide, o progenitores diploides, podrían deberse a que la partenocarpia estaba condicionada por los genes dominantes complementarios derivados de los diploides naturales comestibles, que son diversamente heterocigotos en su composición genética para estos genes. Desde el punto de vista del mejoramiento, esta es una consideración importante ya que las plantas partenocárpicas pueden ser detectadas sin esfuerzo en los cruzamientos que involucran estos progenitores diploides. En los cruzamientos AA × AA, se obtuvieron 5 híbridos diploides partenocárpicos pero no triploides o tetraploides partenocárpicos. Sin embargo, esto no puede ser aseverado como una regla, ya que en un estudio anterior con los mismos progenitores se obtuvieron híbridos triploides (Sathiamoorthy 1987), aunque la proporción fue baja. Los híbridos partenocárpicos obtenidos de los cruzamientos AB × AA merecen una evaluación para ser utilizados como progenitores potenciales en los futuros programas de mejoramiento. Referencias Rowe P.R. & D.L. Richardson. 1975. Breeding bananas for disease resistance, quality and yield. Technical Bulletin No. 2. Tropical Agricultural Research Service, La Lima, Honduras. Simmonds N.W. 1952. Experiments on the pollination of seeded diploid bananas. Journal of Genetics 51: 32-40. Simmonds N.W. 1953. Segregation in some diploid bananas. Journal of Genetics 51:458-469. Simmonds N.W. 1959. Experiments on the germination of banana seeds. Tropical Agriculture Trinidad 29: 259–273. Sathiamoorthy S. 1987. Studies on male breeding potential and certain aspects of breeding bananas. Ph.D. Thesis, Tamil Nadu Agriculture University, Coimbatore. Efectos del desmane y la distancia de siembra sobre las características productivas del plátano ‘FHIA-20’ E InfoMusa - Vol. 13 - No.1 Los autores trabajan en el Department of Fruit Crops, Horticultural College and Research Institute, Tamil Nadu Agricultural University, Coimbatore-641 003, Tamil Nadu, India. La dirección actual de V. Krishnamoorthy es Krishi Vigyan Kendra, Vriddhachalam - 606 001, Cuddalore, Tamil Nadu, India, e-mail: [email protected] Prácticas culturales M. Aristizábal L. n Colombia la comercialización del plátano se realiza por racimos, pero últimamente los mercados especializados lo presentan en manos y dedos (Arcila et al. 2000a). En este caso, el tamaño de estos determina el precio de venta (Giraldo 1998). La comercialización por racimos aumenta la vida en anaquel pero su manejo es difícil y origina grandes pérdidas comerciales, al contrario de la comercialización en unidades o manos (Arcila et al. 2000a). En general, las últimas manos son de menor tamaño y se desechan o se venden a menor precio ya que Frutas del híbrido H-201 no cumplen los estándares de calidad de los mercados especializados. El desmane consiste en eliminar varias manos distales (Rodríguez et al. 1988), para que la materia seca no utilizable comercialmente se distribuya entre las manos que permanecen en el racimo, que deben aumentar su tamaño. No se conoce si dicha respuesta puede ser alterada por la distancia de siembra. Como el tamaño del fruto es una característica importante para los mercados especializados, el objetivo de este estudio fue 9 determinar el efecto de la eliminación selectiva de las manos sobre parámetros de producción y la calidad de los frutos del plátano FHIA-20, en función de la distancia de siembra. Materiales y métodos El estudio se llevó a cabo en la granja Montelindo (propiedad de la Universidad de Caldas), ubicada en la región Santágueda, municipio de Palestina (Caldas), a 5º05’ N y 75º40’ W, a 1050 msnm, con temperatura media de 22.5ºC, humedad relativa del 76%, precipitación anual de 2100 mm y brillo solar anual de 2010 horas. En hoyos de 40 cm x 40 cm se sembraron agujas recortadas (500 g, aproximadamente) de plátano FHIA-20, provenientes de la misma granja. Se emplearon distancias de 3 m y 4 m entre surcos por 3 m entre plantas. El manejo agronómico se efectuó oportunamente e incluyó fertilización, deshije, descalcete, deshoje, embolse y manejo de arvenses. Los tratamientos de desmane se efectuaron 15 días después de la emergencia de la bellota. La intensidad de desmane se expresa por el número de manos dejadas en el racimo: seis, siete u ocho manos. No hubo desmane en las plantas testigo pero se les eliminó la bellota. En cosecha se evaluó: peso del racimo, peso promedio del dedo, peso promedio de mano, peso de cada mano, peso promedio del dedo en cada mano y rendimiento. Los datos fueron sometidos a análisis de varianza para un diseño de bloques completamente al azar, en un arreglo factorial de 2x4, con cuatro repeticiones y siete plantas útiles por repetición. La separación de medias de tratamientos se efectuó mediante la prueba de Tukey. Para todos los análisis se empleó el programa SAS (Statistical Analysis System). Resultados Se obtuvieron diferencias altamente significativas para todas las variables, entre los niveles de desmane. El efecto de la distancia de siembra sólo fue altamente significativo para el rendimiento, mientras que el efecto de la interacción entre distancia de siembra y desmane siempre fue significativo (Tabla 1). En ambas distancias, los testigos tuvieron los mayores pesos de racimo, pero los valores más bajos de pesos promedios de dedo y mano, debido a que, en promedio, el número de manos por racimo fue de 11. En racimos desmanados, el peso de racimo aumentó si se dejaban más manos, debido al mayor número de dedos, ya que los pesos promedios de dedo y mano mostraron relación inversa con el numero de manos por racimo (Tabla 2). En racimos con seis manos, el peso de racimo fue el más bajo, pero los pesos promedios de dedo y mano fueron los más altos (Tabla 2). El peso promedio de mano fue 48.7%, 29.3% y 14.6% superior en racimos con seis, siete y ocho manos, respectivamente, en comparación con el testigo a 3 m x 3 m; los valores correspondientes para 4 m x 3 m Tabla 1. Cuadrados medios y niveles de significancia del desmane y de la distancia de siembra sobre los parámetros agronómicos de FHIA-20. Fuente de variación Desmane Distancia Interacción Residual R2 CV GL 3 1 3 24 Peso de racímo 100.7** 8.3 11.6** 3.7 0.821 5.12 Peso promedio de mano Peso promedio de dedo Rendimiento 4.2** 25501** 93.3** 0.16 5832 1100** 0.24* 6104 * 13.5 * 0.06 2011 4.2 0.926 0.722 0.941 5.04 12.14 5.15 *, ** Efectos significativos (5%) o altamente significativos (1%) según la prueba de Fisher. Tabla 2. Efecto del desmane y de la distancia de siembra sobre los parámetros agronómicos de FHIA-20. Distancia de siembra Tratamiento de desmane Peso de racimo (kg) Peso promedio de mano (kg) Peso promedio de dedo (g) Rendimiento (T/ha) 3mx3m 6 manos 7 manos 8 manos Testigo 36.6 cd 37.3 cd 37.5 bcd 44.6 a 6.1 a 5.3 bc 4.7 d 4.1 e 497 a 375 b 342 b 308 b 40.6 bc 41.4 b 41.6 b 49.5 a 6 manos 7 manos 8 manos Testigo Diferencia mínima significativa (5%) 33.1 d 37.7 bcd 39.1 bc 42.0 ab 4.67 5.5 ab 5.4 bc 4.9 cd 3.8 e 0.593 393 b 387 b 357 b 297 b 106.4 27.5 f 31.4 ef 32.6 ed 41.6 b 4.87 4mx3m Promedios dentro de cada columna acompañados de letras distintas denotan diferencias significativas según la prueba de Tukey. 10 InfoMusa - Vol. 13 - No.1 fueron 44.7%, 42.1% y 28.9% indicando que el efecto fue mayor en la distancia más amplia. El rendimiento fue significativamente mayor en la menor distancia, debido a las diferencias de población: 1111 y 833 plantas/ha, para 3 m x 3 m y 4 m x 3 m, respectivamente; no obstante, en la distancia menor las diferencias entre tratamientos de desmane no fueron significativas (Tabla 2). Los promedios para tratamientos de desmane a través de las distancias de siembra indican que, en comparación con el testigo, el peso de racimo se reduce significativamente en 19.4%, 13.4% y 11.5% en racimos con seis, siete y ocho manos, respectivamente. Esta situación también se refleja en el rendimiento, cuyas reducciones fueron de 19.2%, 14.0% y 12.3% respectivamente. Así, en volumen de producción, el desmane resultaría desventajoso para el productor, en especial si los racimos se dejan con seis manos. El peso inferior de racimo se debe más al efecto del número de dedos, cuyos totales fueron 78, 97 y 111 cuando los racimos se dejaron con seis, siete y ocho manos respectivamente, que al tamaño de los mismos, ya que el peso promedio de dedo fue superior en racimos con menos manos. En comparación con el testigo, el peso promedio de dedo se incrementó significativamente en 42.9%, 22.1% y 11.9% cuando los racimos se dejaron con seis, siete y ocho manos, respectivamente, mientras que peso promedio de mano lo hizo, también significativamente, en 48.7%, 35.9% y 23.1%. Así, para la calidad del producto, la mejor respuesta se obtiene con racimos de seis manos. En ambas distancias de siembra, el peso promedio de dedo en cada mano presentó los valores más bajos en el testigo. A 3 m x 3 m, el peso promedio de dedo de la primera mano, en racimos con ocho, siete y seis manos, superó en 21.9%, 48.3% y 52.0%, respectivamente, el valor correspondiente en el testigo; a 4 m x 3 m, los valores fueron 14.8%, 42.6% y 29.5%, respectivamente (Tabla 3). A 3 m x 3 m, el peso promedio de dedo aumentó a medida que los racimos tenían menor número de manos; a 4 m x 3 m dicho comportamiento fue errático. En general, los racimos de plátano tienden a tener forma triangular ya que las manos ubicadas en la zona proximal son de mayor tamaño que las localizadas en la parte distal (Aristizábal 1995). En este estudio, el peso de la última mano equivalió al 28.3% del peso de la primera en racimos testigo en la distancia de 3 m x 3 m, mientras que en la distancia de 4 m x 3 m, dicho valor fue de 21.3%; pero con desmane, las proporciones fueron del 43.5%, 43.0% y 58.8% para racimos con ocho, siete y seis manos, respectivamente, a 3 x 3 m, y de 38.5%, 42.7% y 49.4%, respectivamente, a 4 x 3 m. Así, el desmane reduce el número de dedos y hace que el racimo sea menos triangular. Discusión En el plátano FHIA-20, el desmane reduce el peso del racimo pero mejora la calidad comercial de la producción, ya que aumenta el peso de las manos y los dedos, especialmente cuando el racimo se deja con seis manos. El incremento en el peso de manos y frutos a causa del desmane ha sido reportado por Arcila et al. (2000b) en FHIA-21, Rodríguez et al. (1988) en ‘Superplátano’, ‘Laknau’ y ‘Maricongo’ y por Quintero y Aristizábal (2003) en ‘Dominico hartón’. Pero este efecto no compensó la pérdida de peso del racimo, lo cual coincide con lo reportado por Irizarry et al. (1994). Con el desmane se pretende que la materia seca destinada para el llenado de las manos eliminadas se distribuya en las que quedan en el racimo, lo cual es teóricamente válido si se tiene en cuenta que no hay modificación en el área foliar funcional para el llenado del racimo. De acuerdo con la teoría sobre la relación fuente - receptáculo, el desmane implica disminución del tamaño del receptáculo y reducción en la Tabla 3. Peso promedio (g) de los dedos en racimos de FHIA-20 sometidos a tratamiento de desmane en dos distancias de siembra. Número de la mano 1 2 3 4 5 6 7 8 9 10 11 Distancia de 3 m x 3 m Testigo 333 347 347 343 314 315 285 293 267 270 189 Ocho 406 381 386 343 331 292 283 250 InfoMusa - Vol. 13 - No.1 Siete 494 412 446 357 353 317 309 Distancia de 4 m x 3 m Seis 506 473 478 467 454 427 Testigo 359 341 306 329 315 285 283 264 280 256 186 Ocho 412 406 367 350 338 333 275 245 Siete 512 412 373 371 336 373 350 Seis 465 412 400 350 367 325 11 actividad de la fuente, es decir, en la actividad fotosintética de la hoja (Shibles 1984). Esto explica por qué en plantas con similar número de hojas funcionales en floración (12 en promedio, en el presente estudio), el peso de racimo en racimos desmanados es menor que en racimos intactos. El efecto benéfico del desmane está en que mejora el tamaño comercial de los frutos, especialmente cuando el producto se comercializa en manos o en dedos, o se destina para la venta en mercados especializados, como es la tendencia actual (CCI 2000). En éstos casos, el peso de las manos o de los dedos es un factor determinante para establecer el precio de venta del producto y, en consecuencia, los ingresos del productor. Belalcázar y Cayón (1998), observaron en ‘Dominico hartón’ que el aumento en la distancia de siembra incrementa el peso de racimo. En éste estudio, las mejores respuestas se obtuvieron a 3 x 3 m, lo cual sugiere que las condiciones microclimáticas imperantes en el ambiente formado en tal distanciamiento, particularmente la radiación solar, no ofrecieron limitación para el crecimiento y producción de la planta. Referencias El autor es Profesor Titular, Departamento de Fitotecnia, Universidad de Caldas, PO Box 275, Manizales, Colombia. Prácticas culturales Arcila P.M.I., G. Giraldo, C.S. Belalcázar, G. Cayón & J.C. Méndez. 2000a. Comportamiento postcosecha de los plátanos Dominico Hartón y FHIA-21 en diferentes presentaciones. Pp. 79-84 in Postcosecha y Agroindustria del Plátano en el Eje Cafetero de Colombia (D.G. Cayón Salinas, G.A. Giraldo Giraldo & M.I. Arcila Pulgarín, eds). CORPOICA y Universidad del Quindío. Armenia, Colombia. Arcila P.M.I., J.A. Valencia, C.S. Belalcázar & H. Morales. 2000b. Efecto del desmane sobre la calidad y la producción del híbrido de plátano FHIA-21. Pp. 7178 in Postcosecha y Agroindustria del Plátano en el Eje Cafetero de Colombia (D.G. Cayón Salinas, G.A. Giraldo Giraldo & M.I. Arcila Pulgarín, eds). CORPOICA y Universidad del Quindío. Armenia, Colombia. Aristizábal L.M. 1995. Componentes del rendimiento en plátano (Musa AAB cv Dominico hartón). Revista Universidad de Caldas 15(1-2):65-75. Belalcázar C.S. & D.G. Cayón.1998. Altas densidades de siembra. Pp.147-154 in Seminario Internacional de Producción de Plátano. Memorias. (M.J. Giraldo Cardona, S. Belalcázar Carvajal, D.G. Cayón Salinas & R.G. Botero Izaza, eds). INIBAP, Palmira, Colombia. CCI. 2000. Acuerdo de competitividad de la cadena productiva del plátano en Colombia. Corporación Colombiana Internacional, Secretaría Técnica, Bogotá. Giraldo G.G. 1998. Conservación e industrialización del plátano. Pp. 266-273 in Seminario Internacional de Producción de Plátano. Memorias. (M.J. Giraldo Cardona, S. Belalcázar Carvajal, D.G. Cayón Salinas & R.G. Botero Izaza, eds). INIBAP, Palmira, Colombia. Irizarry H., E. Hernández & J. Rodríguez. 1994. Yield of the dwarf banana cultivars grown with minimum tillage in the Puerto Rico’s mountain region. The Journal of Agriculture of the University of Puerto Rico 78(1-2):1-7. Quintero S.J.A. & L.M. Aristizábal. 2002. Efectos del desmane sobre las características productivas de ‘Dominico hartón’ y ‘África’ en Colombia. INFOMUSA 12(1):44-45. Rodríguez J.A., H. Irizarry & E. Rivera. 1988. Efecto de la poda de manos en el rendimiento y calidad de las frutas del plátano (Musa acuminata x Musa balbisiana, AAB). Pp. 537-541 in Memorias VIII Reunión ACORBAT. Medellín, Colombia. Shibles R. 1984. Crop Physiology. Iowa State University Press. Efecto de la profundidad de plantación sobre la duración del ciclo de cultivo y rendimiento S. B. Bakhiet y G. A. A. Elbadri E xisten dos principales métodos para la siembra de plantas de banano, a saber, en hoyos y en surcos (Simmonds 1966). La profundidad de plantación varía con el tipo de suelo y el tipo de material de plantación (Robinson 1995). Ya que el crecimiento de los retoños usualmente tiene lugar desde las partes centrales y superiores del cormo, existe una tendencia para que los retoños sucesivos nazcan cerca de la superficie del suelo e incluso sobre la superficie del suelo (Simmonds 1966). En las plantas establecidas a partir de retoños, el sistema radical es adventicio y las condiciones desfavorables aumentan la sensibilidad de la planta al estrés hídrico. La reducción de la extensión del sistema radical resulta en un menor y menos seguro anclaje de las plantas. Estas plantas están propensas a volcarse bajo el peso 12 de un racimo prematuro, especialmente durante las estaciones con fuertes vientos o lluviosas. La mata entera puede desarraigarse, dejando el área improductiva para la vida de la plantación. En la región de Kassala, Sudan, las plantas de banano usualmente son sembradas a una profundidad de 30 a 40 cm. Una alta proporción de las plantaciones de banano se convierte en plantaciones improductivas después del cuarto ciclo de cultivo. La reducción en el rendimiento se podría atribuir a profundidades de siembra relativamente superficiales. Varios trabajadores investigaron la disminución en la producción de los bananos después de unos pocos ciclos de cultivo. Turner (1970) postuló que los cultivos procedentes de los retoños son potencialmente más productivos, ya que se benefician de los residuos de los cultivos anteriores y que la InfoMusa - Vol. 13 - No.1 disminución del rendimiento está asociada con la infección por plagas y enfermedades, particularmente nematodos. Robinson (1995) reportó que los hoyos poco profundos podrían causar una sequía de la planta e inducir un sistema radical superficial en las plantas madre y retoños. Este estudio fue iniciado para examinar el efecto de la profundidad de plantación sobre la duración del ciclo de cultivo y el rendimiento de los bananos. Materiales y métodos El material de plantación utilizado en esta investigación fueron retoños de espada. Estos fueron sembrados en hoyos 30 cm x 30 cm de diferentes profundidades: 30, 40, 50 y 60 cm. Luego, los hoyos fueron llenados con la tierra extraída. El suelo en esta región es un profundo depósito de limo del área del río Gash. Este tipo de suelo, con una textura de franco limosa a franco limoso arcillosa, tiene la capacidad más alta de humedad disponible del suelo del delta del río Gash (Dijkshoorn 1994). No es ni suelto ni compacto y su perfil típico cubre desde los 40 cm a más de 2 m. Se utilizó un diseño aleatorio de bloques completos con cuatro réplicas y cuatro plantas por parcela. El espaciado de las plantas fue 3 m dentro y entre las filas. La plantación se realizó el 31 de marzo de 1997 en la Kassala Experimental Research Station. Las prácticas culturales, exceptuando la fertilización, se realizaron a medida que se requirieron durante todo el período experimental. Se registró la cantidad de días que se necesitaron para que el cormo empezara a germinar. Cuatro meses después de la siembra se contó la cantidad de retoños nuevos por mata. Se registró la cantidad de días desde la plantación hasta la emisión de la inflorescencia (floración), desde la floración hasta la cosecha y desde cosecha a cosecha durante cuatro ciclos de cultivo. La maduración del racimo fue determinada midiendo el diámetro de la fruta central exterior de la segunda mano durante la cosecha. Los racimos cosechados fueron pesados utilizando una balanza de resortes. Se contó la cantidad de manos por racimo y de dedos por mano. Los promedios fueron comparados utilizando la Prueba de rangos múltiples de Duncan con una probabilidad de 0.05. Resultados y discusión Como se muestra en la Tabla 1, el período para la germinación del cormo y la cantidad de retoños por mata no diferían estadísticamente con las profundidades de plantación. Esto podría ser el resultado de utilizar materiales de plantación de retoños espada uniformes. Bakhiet et al. (2003) han observado diferencias en el período de germinación del cormo cuando el tipo de material de plantación difería entre sí. La cantidad de días desde la plantación hasta la brotación y desde la plantación hasta la cosecha en el primer ciclo de cultivo disminuyó significativamente con el aumento de la profundidad de plantación, pero el período desde la brotación hasta la cosecha no difería estadísticamente. Los resultados muestran que una profundidad de plantación de 60 cm redujo significativamente la cantidad de días desde la plantación hasta la brotación y hasta la cosecha, pero no desde la brotación hasta la cosecha. El intervalo entre las cosechas durante los cuatro ciclos de cultivo difería significativamente con la profundidad de plantación. Una profundidad de plantación de 60 cm dio como resultado un intervalo entre las cosechas significativamente más corto. La única excepción fue el cultivo del 4º ciclo a una profundidad de plantación de 50 cm, que difirió significativamente del cultivo del 4º ciclo plantado a 60 cm (Tabla 1). La excavación de hoyos profundos parece apresurar la floración, mientras que la maduración parece ser controlada por la temperatura durante el desarrollo de los racimos, como lo observó Robinson (1981), entre tanto Fraser y Eckstein (1998) reportaron una tendencia hacia un ciclo más largo como resultado de la plantación profunda utilizando plántulas de banano provenientes del cultivo de tejidos. Esta discrepancia podría ser una consecuencia de la utilización de diferentes materiales de plantación. Los resultados también muestran que el peso del racimo aumentó con la profundidad de plantación, y los racimos más grandes se observaron a 60 cm, con excepción del segundo ciclo de cultivo, donde el peso del racimo a profundidades de plantación de 50 y 60 cm no difirieron significativamente (Tabla 2). Estos descubrimientos están en concordancia con los reportados por Manica (1976), Obiefuna (1983) y Fraser y Eckstein (1998), quienes Tabla 1. Efecto de la profundidad de plantación sobre el desempeño agronómico Profundidad de plantación 30 40 50 60 Tiempo hasta germinación del cormo (días) 29 a* 32 a 32 a 32 a Número de retoños 4 MDS** 2.7 a 3.0 a 2.5 a 2.3 a Tiempo desde la plantación hasta la brotación (días) 314 a 312 a 305 ab 285 b Tiempo desde la brotacíón hasta la cosecha (días) 98 a 95 a 100 a 100 a Tiempo desde la plantación hasta la cosecha (días) 412 a 407 a 405 a 385 b * Los promedios seguidos por la misma letra no difieren significativamente a p= 0.05 de acuerdo a la Prueba de Rangos Múltiples de Duncan. ** MDS: meses después de la siembra; R1 = Segundo ciclo de cultivo; R2 = Tercer ciclo de cultivo. R3 = Cuarto ciclo de cultivo. InfoMusa - Vol. 13 - No.1 13 R1 147 a 136 a 144 a 44 b Tiempo de cosecha a cosecha R2 127 a 128 a 124 a 50 b R3 147 a 125 ab 73 bc 54 c Tabla 2. Efecto de la profundidad de la plantación sobre los parámetros de rendimiento. Prof. (cm) 30 40 50 60 Peso del racimo (kg) MP R1 R2 R3 14.5 b* 16.9 b 16.0 b 15.4 b 14.5 b 16.7 b 16.0 b 16.2 b 15.3 b 19.3 ab 18.6 b 20.0 b 18.2 a 22.3 a 25.5 a 25.1 a Cantidad de manos/racimo MP R1 R2 R3 8.0 c 8.9 b 8.5 b 8.0 c 8.8 b 8.7 b 8.4 b 8.3 bc 8.4 b 9.2 ab 9.1 b 9.6 ab 9.5 a 9.9 a 10.8 a 10.9 a Cantidad de dedos/mano MP R1 R2 R3 17.2 a 18.0 a 18.2 a 17.2 a 17.1 a 17.5 a 17.9 a 18.4 a 17.6 a 18.9 a 17.7 a 18.3 a 18.0 a 19.7 a 18.4 a 18.9 a * Los promedios dentro de la misma columna que tienen la misma letra no difieren significativamente a p= 0.05 de acuerdo a la Prueba de Rangos Múltiples de Duncan. PM = Primero ciclo de cultivo (Planta madre) ; R1 = Segundo ciclo de cultivo; R2 = Tercer ciclo de cultivo; R3 = Cuarto ciclo de cultivo. S. B. Bakhiet trabaja en la Kassala Research Station y G. A. A. Elbadri en la Gezira Research Station, Agricultural Research and Technology Corporation, Wad Medani, P.O. 126, Sudan. Sistema radical observaron que la plantación poco profunda estaba asociada con racimos más pequeños. Los resultados también muestran, que a una profundidad de plantación de 60 cm, el peso de los racimos aumentó hasta el tercer ciclo de cultivo antes de nivelarse (Tabla 2). Esto no fue observado con otros tratamientos en los cuales el peso de los racimos disminuyó después del segundo ciclo de cultivo. Usualmente, el peso del racimo aumenta hasta el cuarto ciclo de cultivo y luego se nivela, con la excepción de cuando existen problemas con el crecimiento de las raíces debido al pobre drenaje o a nematodos (Stover y Simmonds 1987). Los resultados también muestran un efecto significativo de la profundidad de plantación sobre el número de manos por racimo en los cuatro ciclos de cultivo (Tabla 2). La cantidad de manos por racimo de las plantas sembradas en los hoyos de 60 cm difería significativamente de la cantidad de manos en todos los otros tratamientos, con excepción del segundo y cuarto ciclos de cultivo de aquellos plantados en los hoyos de 50 cm. La tendencia fue similar a la observada con el peso de los racimos, dadas las relaciones entre el peso del racimo y la cantidad de manos por racimo. Sin embargo, la cantidad de dedos por mano no varió con la profundidad de plantación (Tabla 2). Aunque una profundidad de plantación de 60 cm parece reducir la duración del ciclo de cultivo y aumentar el peso de los racimos, es necesario realizar más investigaciones para determinar las relaciones entre la profundidad de la plantación, contenido de humedad en el suelo, absorción de nutrientes e infección con los nematodos. Referencias Bakhiet S.B., M.A. Ali & G.A.A. Elbadri. 2003. Effect of planting material on crop cycle duration, growth and yield of banana (Musa AAA group). A paper submitted to the Crop Husbandry Release Committee, Wad Medani, Sudan. Dijkshoorn J.A. 1994. Soil-water-plant relationship in Gash Delta assessment on land-use. Technical Report. Kassala, Sudan. Franser C. & K. Eckstein. 1998. Plantlet size and planting method for tissue culture banana plant. Acta Hort. (ISHS) 490:159-166. Manica I. 1976. The effect of hole size and planting depth on the productivity of the banana cv. Nanicão. Revista Ceres 23:426-429. Obiefuna J.C. 1983. Effect of propagule type and depth of planting on the yield of plantain (Musa AAB) in the rain forest of Nigeria. Tropical Agric. (Trinidad) 60:107-110. Robinson J.C. 1981. Studies on the phenology and production of Williams banana in subtropical climate. Subtropica 2: 12-16. Robinson J.C. 1995. System of cultivation and management. Pp. 15-65 in Bananas and Plantains. (S. Gowen, ed.). Chapman and Hall. Simmonds N.W. 1966. Bananas. Longman, London. Stover R.H. & N.W. Simmonds. 1987. Bananas. Longman, London. Turner D.W. 1970. The growth of the banana. J. Aust. Inst. Agric. Sci. 36:102-111 Relaciones entre la capacitancia eléctrica y características de las raíces G. Blomme, I. Blanckaert, A. Tenkouano y R. Swennen D ada la importancia de las raíces en la absorción de nutrientes y agua, y la estabilidad de la planta, se han realizado muchos esfuerzos en los últimos años para investigar el sistema radical de Musa. Sin embargo, la excavación de las plantas enteras no sólo requiere mucho trabajo, consume tiempo y es destructivo, las características estimadas del sistema radical son válidas solo para un período específico. Tomar muestras representativas de las raíces es una opción (Blomme 2000). Alternativamente, 14 múltiples ecuaciones de regresión pueden tomar ventaja de las fuertes relaciones entre las características de los retoños y las características del sistema radical (Blomme 2000, Blomme et al. 2001). Sin embargo, este método puede ser utilizado en las plantas que se encuentran en la etapa vegetativa. Un método de evaluación rápido y no destructivo que ha sido probado en otras especies de plantas (por ejemplo, zanahorias, maíz, avena, cebolla, girasol y tomate) es la utilización de capacitancia eléctrica (Chloupek InfoMusa - Vol. 13 - No.1 1972, Chloupek 1977, Dalton 1995, Van Beem et al. 1998). Un condensador consiste de dos placas planas paralelas separadas por el vacío o cualquier otro aislador. Una de las placas está conectada a una batería y la otra, a la tierra. Cuando se aplica el voltaje a la primera placa, a la segunda se induce una carga simultánea. Cualquier diferencia en el potencial entre las dos placas es una medición de la capacitancia, C. La capacitancia es dependiente de la constante dieléctrica εr del material no conductor entre las dos placas: C=[εr x A]/d (A=área de la superficie y d=distancia entre las placas). En las plantas, se asume que una corriente eléctrica aplicada a la planta pasará a través de los tejidos vasculares que transportan los nutrientes y agua, y a través del suelo e interface suelo-raíces, sin mayor resistencia. En una planta de banano, el suelo y el tejido vascular de las raíces están separados por una zona de resistencia significativamente más alta, es decir, la corteza da la raíz. Cada segmento de raíz puede representado esquemáticamente como un condensador cilíndrico simétrico que consiste del tejido vascular rodeado por la corteza de la raíz (Figura 1). El borde exterior de la corteza está en contacto con la solución electrolítica del suelo. Las capas central y exterior están separadas por el tejido de corteza menos conductivo. La capacitancia nunca ha sido utilizado antes con los bananos y plátanos, que es sorprendente dado que los tejidos de banano contienen mucha agua. El objetivo de este estudio fue evaluar la posibilidad de utilizar las mediciones de la capacitancia para proporcionar la información sobre el sistema radical del banano. Materiales y métodos Los experimentos se realizaron entre agosto y finales de septiembre de 1999 en la estación High Rainfall del Instituto Internacional de Agricultura Tropical (IITA) en Onne, Nigeria. Los campos experimentales se establecieron utilizando plantas derivadas de los retoños. �r Suelo Corteza Antes de la siembra, el suelo fue arado superficialmente. El diseño experimental fue un diseño de parcela dividida dentro de un diseño de bloque completamente al azar con dos réplicas de dos plantas derivadas de los retoños por genotipo. El principal tratamiento de la parcela fue el tiempo de observación (24, 29 y 33 semanas después de la plantación). El segundo tratamiento fue el genotipo: ‘Mbi egome’ (AAB) y ‘Fougamou’ (ABB). El espaciado fue de 3.5 m x 3.5 m para prevenir la superposición de los sistemas radicales adyacentes. Todas las medidas de la capacitancia se realizaron con un medidor de capacitancia portátil, tipo ISO-TECH 9023 RS 205-7496. Basándose en las pruebas preliminares, la escala fue fijada en 2µF (10-6 F) a una frecuencia de 800 Hz. Ambos polos fueron unidos a los electrodos de aguja de cobre utilizando pinzas de cocodrilo. Un electrodo positivo de 45 cm de largo fue insertado en el suelo. La corriente pasaba a través del tejido de la corteza, llegaba al cilindro central, es decir, el tejido vascular de la raíz, continuaba a través del tejido vascular del cormo y llegaba al electrodo negativo de 15 cm de largo insertado en el pseudotallo. El circuito se cerraba con un medidor de capacitancia (Figura 2). El electrodo negativo se colocó a dos alturas en el pseudotallo (0 y 10 cm) y a dos profundidades en el pseudotallo (1.5 y 8 cm), y el electrodo positivo se posicionó a diferentes distancias desde el pseudotallo (40, 80 y 120 cm) y profundidades en el suelo (10, 20, 30 y 40 cm). Las mediciones se realizaron en cada uno de los cuatro cuadrantes alrededor de la planta madre. La línea 0º se trazó desde la planta madre hasta el retoño más alto. Los cuadrantes se evaluaron en el sentido de las agujas del reloj, donde el primer cuadrante representaba el área de 90° a la izquierda de la línea 0°. El valor de la capacitancia, que varía con el tiempo, fue observado 60 segundos después de encender el medidor. Todas las mediciones de capacitancia se realizaban dos horas después de la lluvia o riego. DP metro HP D DS Cilindro central Suelo Figura 1. Un segmento de la raíz visualizado como un condensador cilíndrico. El cilindro central y el suelo están separados por la corteza de la raíz menos conductiva cuya constante dieléctrica es εr InfoMusa - Vol. 13 - No.1 Figura 2. Características que definen la posición de ambos electrodos (HP: Altura en el pseudotallo, DP: Profundidad en el pseudotallo, D: Distancia desde el pseudotallo, DS: Profundidad en el suelo) 15 Basándose en un experimento realizado por Chloupek (1977), se investigó un método alternativo para medir la capacitancia. Cuatro electrodos se posicionaron en un cuadrado alrededor de la planta madre e interconectados por un alambre de cobre fino. La pinza de cocodrilo se sujetó al electrodo en la posición de 0º, es decir, la posición del retoño más alto. Las mediciones se realizaron a las distancias de 40 y 80 cm desde la planta madre. La profundidad del suelo permaneció constante a 40 cm. Las posiciones para el electrodo negativo fueron las mismas que en el método estándar. Las mediciones se realizaron en todas las plantas en ambos campos experimentales. Debido a la presencia de un cormo subterráneo, se llevó a cabo un experimento adicional para determinar el efecto del tamaño del cormo sobre las mediciones de capacitancia. Se excavaron diez cormos de tres genotipos (FHIA-03, TMPx548-9 y TMPx1658-4). Se seleccionaron los cormos con dimensiones diferentes. Se cortaron todas las raíces y el pseudotallo. La capacitancia de cada cormo fue medida mientras el cormo estaba parcialmente sumergido en un tanque con agua. Un electrodo fue insertado a una profundidad de 1 cm en el cormo, sin que hiciera el contacto con el agua. El otro electrodo fue mantenido en el agua, a una distancia constante de 20 cm desde el cormo flotante. Después de realizar las mediciones de capacitancia, cada cormo fue cortado por la mitad y se evaluaron la altura y el ancho del cormo. En cuanto se completaron las mediciones de capacitancia, cada planta fue excavada cuidadosamente y se evaluaron las siguientes características del retoño y de las raíces: altura de la planta, circunferencia de la planta, cantidad de retoños, altura del retoño más alto, área foliar de la planta madre, peso del cormo de la planta madre, peso del cormo de los retoños, cantidad de raíces adventicias de la mata, peso seco de las raíces de la mata, largo de las raíces adventicias de la mata y el diámetro promedio de 40 raíces adventicias seleccionadas al azar. El área foliar fue calculada de la siguiente manera: largo de la hoja x el ancho más grande de la hoja x 0.8 (Obiefuna y Ndubizu 1979). El largo de las raíces adventicias fue medido utilizando el método de cruzamiento de las líneas (Tennant 1975). El método de cruzamiento de las líneas consiste en esparcir las raíces adventicias en una rejilla y contar la cantidad de puntos de cruzamiento, que luego se multiplican por el factor de conversión 2.3571, apropiado para una rejilla de 3 cm x 3 cm. El diámetro basal de las raíces adventicias se midió utilizando un calibrador Vernier. Se realizó un análisis de varianza utilizando el paquete estadístico SAS. Tomando en cuenta 16 todos los datos, se realizaron los análisis de diagrama de dispersión y correlación lineal (Proc CORR en SAS), pero también tomando en cuenta la edad de las plantas. Resultados Se descubrieron correlaciones poco significativas entre los valores de capacitancia y características del sistema radical para todas las combinaciones de las posiciones de los electrodos y etapas de desarrollo de las plantas (no se presentan los datos). Solo el 12.5%, 8% y 5.7% de las correlaciones entre la capacitancia y la cantidad de las raíces adventicias, peso seco de las raíces y el largo de las raíces adventicias, respectivamente, fueron significativos. Ninguna posición puntual dio correlaciones más significativas y las correlaciones poco significativas entre las características del sistema radical y los valores de capacitancia no estaban consistentemente positivos. Finalmente, ninguno de los cuatro cuadrantes evaluados dio un mayor número de correlaciones positivas. Aunque la capacitancia no estaba correlacionada significativamente con ninguna característica del sistema radical, la posición del electrodo afectó las lecturas de capacitancia. La distancia del electrodo desde el pseudotallo no tuvo efecto significativo sobre los valores de capacitancia, resultado que concuerda con las observaciones realizadas en el maíz por Van Beem et al. (1998), pero su altura y la profundidad en el pseudotallo y su profundidad en el suelo si tuvieron efecto (Tabla 1). El aumento de la altura a la cual el electrodo estaba insertado causó una caída en la capacitancia ya que una mayor cantidad del tejido de la planta en el circuito eléctrico aumentó la resistencia en este circuito. Por otro lado, el aumento de la profundidad a la cual se insertó el electrodo en el pseudotallo aumentó la capacitancia. Los valores más altos de la capacitancia se obtuvieron cuando el electrodo estaba insertado a mayores profundidades en el suelo. El cuadrante también tuvo un efecto significativo sobre los valores de capacitancia (Tabla 2). El efecto significativo del cuadrante puede indicar una influencia de la distribución desigual de las raíces y tejido del cormo alrededor de la planta madre. Ya que el lado en el cual se desarrolla el retoño tendría una mayor densidad de las raíces, se esperaba que los valores de capacitancia en este lado serían más altos que en el lado donde no crecían retoños, o sólo crecía uno pequeño. Sin embargo, los resultados muestran que el tamaño del retoño estaba correlacionado negativamente con la capacitancia. El cormo del retoño y, lo que es más importante, los cambios fisiológicos en tiempo en la conexión orgánica entre el retoño y la planta madre, InfoMusa - Vol. 13 - No.1 pueden ser importantes para influenciar los valores de capacitancia. Los valores de capacitancia obtenidos a través del método de cuatro electrodos del suelo interconectados fueron muy correlacionados con los valores obtenidos utilizando el método estándar (R2= 0.87, p<0.0001), en concordancia con las observaciones realizadas por Chloupek (1977) en Helianthus annuus L. cv. ‘Maják’. No se encontraron correlaciones significativas entre la altura y el ancho del cormo y los valores de capacitancia (Tabla 3). Contrario a los cultivos evaluados en investigaciones anteriores con respecto a la capacitancia, las plantas de banano forman numerosos cormos al nivel del suelo. Estos cormos pueden impedir que el circuito eléctrico de un cuadro correcto del sistema radical. Discusión Nuestros resultados muestran que la medición de la capacitancia no puede ser utilizada para estimar las características de las raíces de las plantas jóvenes y adultas de Musa en el campo. La falta de correlación entre los valores de capacitancia y las características de las raíces puede estar relacionada con la morfología de la planta de banano. Por ejemplo, la posición de los haces vasculares puede ser significativa. El electrodo no estaba conectado con el tallo real de la planta (que es el cormo) pero en cambio estaba insertado en el pseudotallo, que consiste de vainas foliares. Este tejido contiene cavidades de aire, que pueden afectar la conductividad, pero no se conoce la magnitud hasta la cual estos factores afecten los resultados. Otra característica de la planta de banano es la presencia del cormo subterráneo y la formación de los retoños. La presencia, posición y dimensiones de los retoños, en particular, parecen ser significativas. Aunque las evaluaciones en el campo se realizaron bajo condiciones de riego y la copa foliar atenuó las fluctuaciones extremas de la temperatura del suelo, pudieron haber ocurrido cambios en la temperatura del suelo que podrían influir sobre las mediciones de capacitancia. En adición, aunque el suelo fue arado antes de la plantación, aún pudieran existir en el suelo cambios estructurales abruptos y cavidades que podrían haber influenciado el circuito eléctrico. Las investigaciones anteriores (Van Beem et al. 1998, Dalton 1995, Kendall et al. 1982, Chloupek 1977) identificaron la humedad y la temperatura del suelo como factores clave que influyen sobre la capacitancia del sistema radical. La humedad del suelo es importante no solo para la conductividad del suelo pero también para el contacto entre las raíces y el suelo. La capacitancia es solo el reflejo de InfoMusa - Vol. 13 - No.1 Tabla 1. Pruebas de quadrado medio y significación para el valor de capacitancia de ‘Fougamou’ y ‘Mbi Egome’ (4 plantas por cultivar, 24 semanas después de la plantación). Fuente de variación Altura en el pseudotallo Profundidad en el pseudotallo Distancia desde el pseudotallo Profundidad en el suelo Df 1 2 2 3 Fougamou 0.32515392*** 0.78452294**** 0.00573300 0.59036384*** Mbi egome 0.43843809*** 0.57899544*** 0.01609219 0.88199040*** Tabla 2. Pruebas de quadrado medio y significatión para el valor de capacitancia (2 cultivares ‘Fougamou’ y ‘Mbi Egome’, 4 plantas por cultivar, 24 semanas después de la plantación. Fuente de variación Cuadrante Altura en el pseudotallo Profundidad en el pseudotallo Distancia desde el pseudotallo Df 3 1 1 1 0.17668901*** 1.07109907*** 0.13158450* 0.04201376 Tabla 3. Coeficiente de correlación entre los valores de capacitancia y caracteristicas del cormo. Altura del cormo Ancho del cormo Valor de capacitancia 0.29 0.38 Altura del cormo 0.91*** las características del sistema radical donde existe un buen contacto eléctrico entre la superficie de las raíces y el suelo. De este modo, una escasez de agua llevará a un bajo valor de capacitancia. Este valor podría ser interpretado falsamente como un indicio de una baja masa radical, mientras que en realidad esto podría ser el reflejo de la baja humedad del suelo. El fuerte efecto de la temperatura del suelo sobre la capacitancia desempeña un papel importante para la interpretación de los resultados y por lo tanto puede considerarse como un disturbio importante para la medición de la capacitancia en el campo. Se asume que un aumento en la temperatura del suelo aumenta la resistencia del suelo, dando como resultado un valor de capacitancia más bajo. Finalmente, la concentración de los electrolitos en el suelo también afecta a la conductividad. A medida que crece la planta, los nutrientes se absorben del suelo, resultando en cambios espaciales en condición nutricional del suelo que también podrían afectar las mediciones de la capacitancia. Agradecimientos Se agradece profundamente el apoyo financiero de la Asociación Flamenca para la Cooperación en Desarrollo y Asistencia Técnica (VVOB) y de la Dirección General Belga para el Desarrollo y Cooperación (DGCD). Referencias Blomme G. 2000. The interdependence of root and shoot development in banana (Musa spp.) under field conditions and the influence of different biophysical factors on this relationship. PhD thesis. 17 G. Blomme trabaja en INIBAP-ESA, P.O. Box 24384, Kampala, Uganda, e-mail: [email protected] pero el trabajo actual fue realizado en el Instituto Internacional de Agricultura Tropical (IITA), High Rainfall Station, PMB 008 NchiaEleme, Rivers State, Nigeria. I. Blanckaert y R. Swennen trabajan en el Laboratorio de Mejoramiento de Cultivos Tropicales, Katholieke Universiteit Leuven, Kasteelpark Arenberg 13, 3001 Heverlee, Bélgica, e-mail: rony.swen [email protected] y A. Tenkouano trabaja en el Centro Ecorregional de Bosques Húmedos, IITA, BP 2008 Messa, Yaoundé, Camerún, e-mail: [email protected]. Dissertationes de Agricultura No. 421. KULeuven, Faculteit Landbouwkundige en Toegepaste Biologische Wetenschappen. Belgium. 183pp. Blomme G., R. Swennen, A. Tenkouano, R. Ortiz & D. Vuylsteke. 2001. Estimación del desarrollo de las raíces a partir de los caracteres de los brotes en banano y plátano (Musa spp.) InfoMusa 10(1):15-17. Chloupek O. 1972. The relationship between electrical capacitance and some other parameters of plant roots. Biol. Plant. 14(3):227-230. Chloupek O. 1977. Evaluation of the size of a plant’s root system using its electrical capacitance. Plant Soil 48: 525-532. Cultivos intercalados Dalton F.N. 1995. In situ root extent measurements by electrical capacitance methods. Plant Soil 173:157-165. Kendall W.A., G.A. Pederson & R.R. Hill. 1982. Root size estimates of red clover and alfalfa based on electrical capacitance and root diameter measurements. Grass Forage Sci. 37:253-256. Obiefuna J.C. & T.O.C. Ndubizu. 1979. Estimating leaf area of plantain. Sci. Hortic. 11:31-36. Tennant D. 1975. A test of a modified line intersect method for estimating root length. J. Ecol. 63:995-1001. Van Beem J, M.E. Smith & R.W. Zobel. 1998. Estimating root mass in maize using a portable capacitance meter. Agron. J. 90:566-570. Productividad del plátano typo False horn intercalado con fríjol de vaca y maíz en el sudeste de Nigeria J.O. Shiyam, B.F. D. Oko y W. B. Binang E l cultivo intercalado de plátano ha sido reportado en la región de los Andes de América del Sur (Stover y Simmonds 1987), Antillas (Rao y Edmund 1984) y Filipinas (Alviar y Cuevas 1976). En los bosques húmedos de Africa Occidental, Devos y Wilson (1979), Ikeorgu et al. (1989) y Oko et al. (2000) han observado que el cultivo intercalado es practicado usualmente por familias con bajos ingresos para maximizar el uso de la tierra y proporcionarse comida y dinero adicionales. En los trópicos húmedos, especialmente en el sudeste de Nigeria, los plátanos se intercalan frecuentemente con cultivos alimentarios como el maíz, la colocasia y vegetales (Francis et al. 1976). El cultivo intercalado de plátano por parte de los agricultores con escasos recursos también ha sido reportado por Karikari (1972) y Obiefuna (1984). Pero el cultivo de plátano usualmente es rentable por uno o dos años después de los cuales la fertilidad del suelo declina resultando en rendimientos anuales bajos de 4 a 8 toneladas por hectárea (Nweke et al. 1988), en comparación con 30 a 50 toneladas por hectárea observadas en los jardines traseros, donde el suelo puede sostener rendimientos altos durante muchos años (Wilson 1987). A pesar de la popularidad del cultivo intercalado de plátano, pocos estudios experimentales de este sistema de cultivo se han realizado en Nigeria. En consecuencia, poco se conoce sobre las interacciones entre las plantas, el rendimiento y la productividad total de los sistemas de cultivo intercalado en los cuales el plátano es el componente principal, probablemente debido a la dificultad de evaluar los rendimientos de los sistemas de cultivo tradicionales. Este ensayo fue diseñado para determinar el efecto sobre la productividad del plátano intercalado con el fríjol de vaca o el maíz. 18 Materiales y métodos El experimento fue conducido durante las estaciones de cultivo en 1998 y 1999 en la finca de enseñanza e investigaciones de la Universidad de Calabar en el cinturón de bosques húmedos en el sudeste de Nigeria. El clima del área es húmedo y soporta una vegetación de bosque húmedo tropical de tierras bajas. La precipitación anual es de alrededor de 2000 a 3000 mm, la mayoría de ella cae entre los meses de marzo y noviembre. La precipitación muestra una distribución bimodal característica, con el máximo en julio y septiembre. Las temperaturas máxima y mínima son de 33°C y 23°C, respectivamente, mientras que la humedad relativa promedio es de 80%. El sitio experimental se encuentra en un suelo franco arenoso (Cobbina et al 1990) con un pH de 5.8, contenido de carbón orgánico de 2%, contenido de N total de 0.17%, índice de fósforo (Bray P1) de 55.7 ppm, acidez intercambiable de 1.28, capacidad de intercambio de cationes (CEC) de 4.14 meq/100g y contenido de magnesio de 0.6 meq/100g. Un plátano Falso Cuerno (False horn), conocido comúnmente en Africa Occidental como ‘Agbagba’ fue intercalado con una variedad de fríjol de vaca (Vigna unguiculata WaIp.) (1T82D-719), recomendada para la zona agroecológica de bosque húmedo de Nigeria (Enwezor et al. 1989), y una variedad local de maíz de maduración media (Zea mays, L.) conocida popularmente como ‘Jkom white’. El experimento fue dispuesto en un diseño de bloques completamente al azar con cuatro réplicas. Los tratamientos fueron: solo plátano; solo fríjol de vaca; solo maíz; plátano y fríjol de vaca; plátano y maíz; plátano, fríjol de vaca y maíz. InfoMusa - Vol. 13 - No.1 Los retoños se obtuvieron de fincas viejas del área. Estos fueron plantados a 2 m de distancia en hoyos de 40 cm de profundidad y 30 cm de ancho llenados con tierra. Los cultivos intercalados fueron sembrados entre las filas, inmediatamente después de que el plátano fue sembrado. Las densidades de plantación fueron 1666 plantas/ha para el plátano, 20 000 plantas/ ha para el maíz y 55 000 plantas/ha para el fríjol de vaca. El tamaño de una unidad de parcela fue de 10 m x 3 m y consistió de una fila de seis matas de plátano 2 m aparte en el centro de la parcela y los cultivos intercalados fueron plantados a ambos lados de la fila de plátano. Cada planta de plátano recibió 400 g de N:P:K (20:10:10) tres meses después de la siembra y 300 g seis y nueve meses después de la siembra para un total de 250 kg/ha (Swennen y Wilson 1985). El maíz y el fríjol de vaca recibieron respectivamente una aplicación total de 120 kg/ha y 20 kg/ha de N:P:K (20: 10:10) (Enwezor et aI. 1989). Las parcelas fueron mantenidas libres de malezas mediante deshierbe con azadón cuando era necesario. El maíz fue cosechado después de 112 días, secado en un horno a 60°C durante 24 horas y descascarado manualmente, mientras que las vainas maduras del fríjol de vaca fueron cosechadas, secadas al sol y descascaradas manualmente. Los racimos de plátano fueron cosechados 90 días después de la floración y el rendimiento por hectárea se basó en el peso promedio de los racimos. Se registraron los datos sobre la altura de la planta 7 y 12 meses después de la siembra, circunferencia del pseudotallo 1 y 12 meses después de la siembra, cantidad de hojas 12 meses después de la siembra, cantidad de días hasta la floración, cantidad de manos por racimo, cantidad de dedos por mano y el peso del racimo, que luego fueron analizados utilizando ANOVA y una prueba de rangos múltiples de Duncan. Resultados Inicialmente el cultivo intercalado redujo la altura del plátano. Este efecto fue el más severo en el tratamiento del plátano intercalado con maíz y fríjol de vaca (Tabla 1). Sin embargo, al momento de la floración, la altura de la planta no difería significativamente. Tampoco hubo diferencias significativas entre los tratamientos con respecto a la circunferencia del pseudotallo y la cantidad de hojas. Los plátanos en monocultivo e intercalados con el fríjol de vaca fructificaron significativamente más temprano que aquellos intercalados con el maíz y con el maíz y fríjol de vaca (Tabla 1). Igualmente, la cantidad promedio de dedos por mano, el peso del racimo y el rendimiento fueron similares en los monocultivos de plátano y en los campos de plátano con fríjol de vaca, pero significativamente más altos que en otros tipos de sistemas intercalados (Tabla 1). El cultivo intercalado redujo significa-tivamente los rendimientos del fríjol de vaca y del maíz en 1998 y 1999 (Tabla 2). Los rendimientos fueron más altos en los monocultivos, seguidos por los sistemas intercalados de dos especies y luego por los de tres especies. Discusión El rendimiento en los campos de plátano/fríjol de vaca se comparó favorablemente con el rendimiento del plátano en los monocultivos debido a la pequeña estatura del fríjol de vaca y su habilidad de fijar el nitrógeno. Estas características minimizan la competencia por la luz y el nitrógeno. En contraste, los rendimientos reducidos del plátano observados en los sistemas intercalados que contenían maíz se deben probablemente a los altos requerimientos Tabla 1. Datos agronómicos registrados para los plátanos después de un ciclo de producción bajo diferentes sistemas de cultivo. Sistema de cultivo Altura de la Circunferencia planta (m) de pseudotallo MDS 12 MDS 12 MDS (cm) Solo plátano 1.6 b 3.9 a 40.6 a Plátano/frijol de vaca 1.3 c 3.9 a 41.7 a Plátano/maíz 0.9 d 3.7 a 40.8 a Plátano/frijol de vaca/maíz 2.3 a 3.8 a 43.0 a Cantidad de hojas 12 MDS 13.0 a 12.9 a 12.8 a 13.3 a Cantidad de días hasta la floración 278 a 306 b 300 b 282 a Cantidad de manos/racimo Cantidad de dedos/mano 7.6 a 6.6 a 6.3 a 7.6 a 52 a 38 b 36 b 54 a MDS: meses después de la siembra Los promedios seguidos por letras diferentes difieren significativamente con la probabilidad de 0.05 de acuerdo a la prueba de rangos múltiples de Duncan. Tabla 2. Rendimiento (t/ha) del plátano, fríjol de vaca y maíz bajo varios sistemas de cultivo. Sistema de cultivo Solo plátano Solo fríjol de vaca Solo maíz Plátano/fríjol de vaca Plátano/maíz Plátano/fríjol de vaca/maíz 1998 Fríjol de vaca Maíz Plátano 19.7 a 1384.6 a 1999 Fríjol de vaca 1131 a 4662.4 a 1313.7 a 1275.4 a Maíz 3967.2 b 2923.2 c 4028.8 a 18.7 a 16.3 b 14.8 b 771.5 b 698.4 c 3661.2 b 2045.8 c Los promedios seguidos por letras diferentes difieren significativamente con la probabilidad de 0.05 de acuerdo a la prueba de rangos múltiples de Duncan. InfoMusa - Vol. 13 - No.1 19 Peso del racimo (kg) 11.2 a 9.8 b 7.2 b 11.8 a Rendimiento (t/ha) 18.7 a 16.3 b 14.8 b 19.7 a de nutrientes del maíz. La competencia por los nutrientes debe haber sido especialmente alta en el segundo año, cuando el plátano entró en su fase reproductora, durante la cual el desarrollo de los racimos demanda gran cantidad de nutrientes (Stover y Simmonds 1987, Oko et al. 2000). La reducción inicial en la altura observada en los plátanos se debió probablemente a la competencia con los cultivos asociados. Las diferencias en los períodos de maduración entre el plátano y otros cultivos puede haber ayudado al plátano a recuperarse del estrés inicial. Esta observación es consistente con la de Andrews (1970). Los rendimientos del fríjol de vaca disminuyeron en todos los sistemas intercalados. De acuerdo a Oko et al. (2000), el fríjol de vaca se desempeña mejor bajo poca o ninguna sombra. La reducción en los rendimientos podría atribuirse a la sombra producida por el plátano y el maíz. Hubo más sombra en 1999 que en 1998. En 1998, el rendimiento promedio se redujo en 109 kg/ha en el sistema intercalado de tres especies, en comparación con el monocultivo, y en 433 kg/ha en 1999. Los rendimientos del maíz también se redujeron y la reducción aumentó con la cantidad de especies en el sistema intercalado y con el tiempo. En 1998, el rendimiento promedio se redujo en 1739 kg/ha en el sistema intercalado de tres especies comparado con el monocultivo, y en 1983 kg/ha en 1999. Oko et al. (2000) reportaron que el maíz a menudo sufre más de la competencia por los nutrientes que de otro factor. También es posible que un aumento en las plagas y enfermedades pueda haber contribuido a la reducción del rendimiento en las especies intercaladas. J.O. Shiyam, B.F. D. Oko y W. B. Binang trabajan en el Department of Crops Science, University of Calabar, Calabar, Nigeria. Híbridos mejorados Referencias Alviar N. & S. Cuevas. 1976. Management of selected Coconut intercropping farms in Laguna, Batangas and Cavite. J. Agric. Economics and Development 6: 184-196. Andrews D.J. 1970. Relay and intercropping with sorghum at Samaru Province. Seminar on intercropping. Ford Foundation/JITA/IART, Ibadan, 1970. Cobbina J.A., A.N. Atta-Krah, A.O. Meregini & B. Duguma. 1990. Productivity of some plants grown on acid soil in Southeastern Nigeria. Tropical Grasslands 24:41-45. Devos P. & G.F. Wilson. 1979. Intercropping of Plantains with food crops: Maize, cassava and cocoyams. Fruits 33:169-173. Enwezor W.O., E.J. Udo, N.J. Usoroh, K.A Ayotade, J.A. Adepetu, V.O. Chude & C.I. Udegbe. 1989. Fertilizer use and management practices for crops in Nigeria. Series No.2. FMAWRRD, Lagos. 163pp. Francis C.A., C.A. Plor & S.R. Temple. 1976. Cultivars for intercropping in the tropics. Pp. 235-253 in Multiple cropping edited by Agronomy society of America Special Publication. Ikeorgu J.E.G, H.C. Ezumah & T.A.T. Wahua. 1989. Productivity of species in cassava/maize/okra/egusi melon complex mixtures in Nigeria. Field crops Research 21:1-7. Karikari S.K. 1972. Plantain growing in Ghana. World Crops. 24:22-24. Nweke F.I., J.E. Njoku & G.F. Wilson. 1988. Productivity and limitations of plantain (Musa spp.) Production in compound gardens in Southeast Nigeria. Fruits 43(3): 161-166. Obiefuna J.C. 1984. Effect of Potassium application during the floral stage plantain (Musa AAB). Fertilizer Research 5:315-319. Oko B.F.D., J.O. Shiyam, A.E. Eneji, T. Honna & S. Yamamoto. 2000. Yield of selected food crops under alley-cropping with some hedgerow species in humid tropical south-eastern Nigeria. Japan. J. Trop. Agr. 44(3):167-190. Rao M.M. & J.E. Edmund. 1984. Intercropping banana with food crops: Cowpea, maize and sweet potatoes. Trop. Agric. 60:9-11. Stover R.H. & N.W. Simmonds. 1987. Bananas. Longman Group UK Ltd. 468pp. Swennen R. & G.F. Wilson. 1985. Flemingia congesta as a live cover plant for plantain. Banana Newsletter 8:17-18. Wilson G.F. 1987. Status of bananas and plantains in West Africa. Pp. 29-35 in Banana and plantain breeding strategies (G. Persley and E.A.L. De Langhe, eds). ACIAR proceedings No.21. ACIAR, Canberra, Australia; INIBAP, Montpellier, France. Efecto del deshije sobre la resistencia de FHIA-23 y SH-3436-9 a enfermedades y plagas A. Vargas y M. Guzmán La Fundación Hondureña de Investigación Agrícola (FHIA) ha desarrollado un significativo programa de mejoramiento genético convencional en musáceas, que ha deparado una considerable cantidad de híbridos mejorados (Rowe 1998, Rowe 1999), dentro de los cuales se encuentran los tetraploides de banano (Musa AAAA) FHIA–23 (Highgate x SH-3362) y SH-3436 20 (Highgate x SH-3142). De este último una selección realizada en Cuba originó el SH-3436-9. FHIA-23 posee buenas características agronómicas y organolépticas y además resistencia parcial a la Sigatoka negra y al Mal de Panamá (Fusarium oxysporum f.sp. cubense) (Orjeda et al. 1999, Rivera y Dueñas 2002). Por su parte, el híbrido InfoMusa - Vol. 13 - No.1 SH-3436-9 produce también satisfactoriamente (Alvarez 1997), sus frutos son de buen sabor, tiene resistencia parcial a la Sigatoka negra (Orjeda et al. 1999) y bajo índice reproductivo de Radopholus similis (J. González 1997, INVIT, com. personal). Existe una considerable cantidad de información de diferentes partes del mundo respecto a la reacción a enfermedades en los híbridos de banano desarrollados por la FHIA, pero es muy escasa la información disponible sobre la respuesta de estos a prácticas de manejo de gran relevancia para una óptima producción, como el deshije y su efecto sobre el comportamiento de los híbridos ante enfermedades como la Sigatoka negra. Con un sistema de deshije orientado, por ejemplo, la línea de siembra, así como la densidad original de población, se mantienen durante un mayor lapso de tiempo con poca alteración, contrario a lo que ocurre con el deshije tradicional (Pérez 2000) Tanto FHIA-23 como SH-3436-9 podrían convertirse en un valioso complemento a la producción de bananos provenientes del subgrupo Cavendish cuyos cultivares son más susceptibles a la Sigatoka negra (Guzmán y Romero 1996, Marín et al. 2003). Por ello, el objetivo del presente trabajo consistió en evaluar el potencial agronómico y la reacción a la Sigatoka negra, a los nemátodos y al picudo negro de ambos híbridos cultivados bajo dos sistemas de deshije (orientada vs. tradicional). Materiales y métodos El trabajo experimental se realizó en el Centro de Investigaciones Agrícolas 28 Millas, propiedad de la Corporación Bananera Nacional (CORBANA, S.A.), entre mayo de 1998 y noviembre de 2001, período que contempló tres ciclos de cultivo. El sitio experimental se ubica a 40 msnm, en el cantón de Matina, provincia de Limón, Costa Rica. Durante el período del estudio la precipitación acumulada fue de 3480, 3652, 3847 y 3857 mm, la temperatura promedio mensual de 24.4, 24.2, 22.5 y 24.2°C y la humedad relativa promedio mensual 87, 88, 89 y 89%, durante los años 1998, 1999, 2000 y 2001 respectivamente. El suelo en donde se implementó la prueba estuvo anteriormente sembrado por cuatro años con los híbridos de banano FHIA-01 y FHIA-02 (Musa AAAB). Por textura, el suelo fue clasificado como Franco arcilloso (arena 34%, arcilla 30%, limo 36%), con pH 6.3; acidez extractable 0.1; materia orgánica 2.8%; Ca 26.4.; Mg 10.1; K 0.7 cmol/L y una capacidad de intercambio catiónico efectiva (CICE) de 37.2 cmol/L. Es considerado taxonómicamente como un InfoMusa - Vol. 13 - No.1 Aquertic Eutrudept (inceptisol arcilloso, con limitaciones de drenaje y características vérticas) (E. Serrano 2002, CORBANA, com. personal). Se utilizaron plantas in vitro como material de siembra. Cada híbrido se evaluó bajo dos sistemas de deshije: direccional u orientado y tradicional, constituyéndose un experimento factorial en un diseño experimental de bloques completos al azar con cuatro repeticiones. Cada parcela experimental constó de 70 plantas y la parcela útil de 40 plantas. Las plantas fueron sembradas en el campo a una distancia de 2.75 m en triángulo para una densidad de 1371 plantas/ha. A las 24 semanas se removieron todos los brotes considerados como provenientes de yemas inferiores del cormo. Posteriormente, se seleccionó, en las parcelas con deshije orientado, un hijo de espada sucesor que mantuviera apropiadamente la orientación de la línea de siembra original y se eliminaron los restantes. En las parcelas con deshije tradicional se seleccionó a la floración aquel hijo de sucesión más desarrollado y con mejor posición. En la segunda y tercera generación, en el deshije orientado, se retuvo aquel primer brote emergido que garantizara con mayor fidelidad el mantenimiento de la línea de siembra original. En el deshije tradicional la selección del hijo de sucesión se realizó a la floración y se retuvo el hijo más vigoroso y mejor ubicado con respecto a las unidades de producción vecinas. La fertilización se realizó mediante aplicaciones mensuales de 15:3:31 (N:P205:K2O) a razón de 83 g por planta. Como medida preventiva contra el volcamiento, las plantas fueron aseguradas mediante la utilización de tensores dobles de polipropileno. El embolse de la fruta se llevó a cabo 15 días después de la emisión de la inflorescencia. Dicha labor se realizó en conjunto con la remoción de las dos últimas manos verdaderas. La cosecha se realizó a las 11 semanas después de la floración. Las evaluaciones a la floración y a la cosecha de las variables correspondientes a la severidad de Sigatoka negra, se efectuaron en 10 plantas por parcela utilizando la escala de Stover modificada por Gauhl (1989). Dichas evaluaciones fueron hechas en las mismas plantas durante los tres ciclos de cultivo. Se midieron las siguientes variables: días a la floración, altura del pseudotallo a la floración, grosor del pseudotallo a la floración medido a una altura equivalente al 25% de la altura del pseudotallo, número 21 total de hojas funcionales a la floración y a la cosecha, peso del racimo, número de manos, diámetro y longitud del fruto central de la segunda mano, número de frutos en la segunda mano, diámetro y longitud del fruto central de la última mano, número de frutos en la última mano, hoja más joven con mancha; es la hoja más joven en presentar el primer estadío de mancha de la Sigatoka negra y se caracteriza por ser una mancha negra visible en ambas superficies de la hoja sin presentar halo clorótico (Fouré 1985) y el índice de infección; es una estimación porcentual del área afectada (necrosada) por la Sigatoka negra en la planta (Romero 1994). No se efectuó combate de nemátodos. Las poblaciones de nemátodos y el estado de las raíces se determinaron en el tercer ciclo mediante el muestreo en hijos de sucesión de plantas recién florecidas, según la metodología propuesta por Araya y Cheves (1998). Tampoco se realizó combate de picudos. El coeficiente de infestación y el grado de pudrición en el cormo se determinaron a la cosecha de las plantas del tercer ciclo mediante el método de Villardebo (1973). No se realizó combate químico de la Sigatoka negra. El tejido foliar necrótico o senescente se eliminó mediante labores fitosanitarias como la deshoja o despunte. El análisis estadístico se hizo para cada ciclo por aparte considerando la estructura factorial de híbridos x sistemas de deshije. Con los datos de las 10 plantas evaluadas por parcela se obtuvo un promedio simple y luego se efectuó un análisis de varianza y separación de medias para cada variable. Resultados En ambos híbridos, no hubo diferencias (P>0.10) entre los sistemas de deshije para la totalidad de las variables que se evaluaron a la floración y a la cosecha en ninguno de los tres ciclos de cultivo. La ausencia (P>0.07) de la interacción híbrido x sistema de deshije permitió, para cada ciclo de cultivo, ignorar el sistema de deshije al comparar los híbridos. FHIA-23 llegó a la floración con mayor altura y diámetro del pseudotallo que SH-3436-9, así como con más cantidad de manos y de días a la floración en cada ciclo de cultivo (Tabla 1). FHIA-23 presentó mayor peso de racimo en los dos primeros ciclos, sin embargo no difirió de SH-3436-9 en el tercero. Con excepción del grosor del fruto de la segunda mano en el segundo ciclo, no hubo diferencias entre ambos híbridos en las dimensiones del fruto ni en su cantidad (número de frutos) en la segunda y última mano (Tabla 1). FHIA-23 mostró a la floración mayor cantidad de hojas totales en los dos primeros ciclos de cultivo, sin diferir con SH-3436-9 en el tercer ciclo. A la cosecha, FHIA–23 mantuvo una similar cantidad de hojas que SH-3436-9 en el primer ciclo y mayor número de ellas en los dos ciclos subsiguientes. No hubo diferencias entre ambos híbridos en el índice de severidad de la Sigatoka negra ni en la ubicación de la hoja más joven con mancha, tanto a la floración como a la cosecha (Tabla 2). Independientemente del híbrido y a excepción de las variables hoja más joven con mancha a la cosecha e intervalo entre floración, donde no se encontraron diferencias entre ciclos de producción, se determinaron diferencias entre ciclos para el resto de las variables estudiadas. En forma general, las variables altura de la planta, grosor del pseudotallo, número de manos, número de frutos en la segunda mano y hoja más joven con mancha a la floración, aumentaron con cada ciclo de cultivo. Por el contrario, el índice de infección de la Sigatoka negra disminuyó. Solamente hubo interacción ciclo x híbrido en las variables: peso del racimo, grosor del Tabla 1. Características de los híbridos FHIA-23 y SH-3436-9 durante tres ciclos de cultivo ( n= 8 parcelas con 40 plantas por parcela útil). Hibrido Primer ciclo FHIA-23 SH-3436-9 Prob>F Segundo ciclo FHIA-23 SH-3436-9 Prob>F Tercer ciclo FHIA-23 SH-3436-9 Prob>F Altura del pseudotallo (m) Grosor del Número de pseudo manos por tallo racimo (cm) Días a floracion1 Peso del racimo (kg) Grosor* del fruto 2a mano Largo del fruto 2a mano Número de Grosor* Largo frutos en la del fruto en del fruto en 2a mano la última la última mano mano (cm) Frutos en la última mano 3.3 ± 0.1 2.9 ± 0.1 0.0009 25.8 ± 0.3 22.3 ± 0.5 0.0001 11.3 ± 0.3 9.9 ± 0.3 0.0151 365 ± 10 342 ± 8 0.0556 23.4 ± 1.1 20.2 ± 1.2 0.0437 40.6 ± 0.4 41.4 ± 0.5 0.8838 21.3 ± 0.2 21.6 ± 0.3 0.5430 18.2 ± 0.3 17.9 ± 0.7 0.4697 33.4 ± 0.5 16.0 ± 0.3 35.4 ± 0.4 17.2 ± 0.3 0.8798 0.5553 15.1 ± 0.3 14.4 ± 0.3 0.8918 3.9 ± 0.1 3.4 ± 0.1 0.0001 30.6 ± 0.4 27.5 ± 0.6 0.001 12.8 ± 0.3 11.5 ± 0.3 0.0004 327 ± 7 284 ± 6 0.0001 28.1 ± 0.8 22.7 ± 1.0 0.0006 40.5 ± 0.3 39.1 ± 0.4 0.0461 21.8 ± 0.1 21.3 ± 0.3 0.2867 19.3 ± 0.3 19.0 ± 0.3 0.3735 33.5 ± 0.3 16.2 ± 0.2 33.5 ± 0.4 16.6 ± 0.2 0.9288 0.2845 15.6 ± 0.2 15.3 ± 0.2 0.1613 4.2 ± 0.1 3.7 ± 0.1 0.0001 34.5 ± 0.3 31.3 ± 0.6 0.0007 12.7 ± 0.3 12.0 ± 0.3 0.0654 324 ± 5 290 ± 6 0.0033 22.2 ± 1.0 22.4 ± 1.4 0.8495 37.4 ± 0.5 38.2 ± 0.6 0.3362 19.9 ± 0.3 20.5 ± 0.4 0.2655 19.2 ± 0.3 20.4 ± 0.7 0.1552 31.5 ± 0.2 14.8 ± 0.2 32.3 ± 0.7 15.3 ± 0.2 0.2603 0.1488 15.5 ± 0.2 15.9 ± 0.2 0.2814 * medido en treintaidosavos de pulgada 22 InfoMusa - Vol. 13 - No.1 Tabla 2. Respuesta de los híbridos FHIA-23 y SH-3436-9 a la Sigatoka negra a la floración y a la cosecha durante tres ciclos de cultivo (n=8 parcelas con 10 plantas por parcela útil). Híbrido Primer ciclo FHIA-23 SH-3436-9 Prob>F Segundo ciclo FHIA-23 SH-3436-9 Prob>F Tercer ciclo FHIA-23 SH-3436-9 Prob>F Hojas totales Floración Indice de infección Hoja más joven con mancha Hojas totales Cosecha Indice de infección Hoja más joven con mancha 8.9 ± 0.1 8.7 ± 0.1 0.0515 54.2 ± 0.5 53.0 ± 0.9 0.2367 2.9 ± 0.1 3.1 ± 0.1 0.4749 3.5 ± 0.2 3.5 ± 0.1 0.9996 83.9 ± 0.7 85.0 ± 1.4 0.4846 1.0 ± 0.0 1.0 ± 0.0 0.3559 9.4 ± 0.1 9.1 ± 0.1 0.0191 44.7 ± 1.2 44.6 ± 1.1 0.9022 3.1 ± 0.1 3.1 ± 0.1 0.7453 4.5 ± 0.2 4.0 ± 0.1 0.0314 73.9 ± 0.8 72.7 ± 1.5 0.5468 1.0 ± 0.0 1.0 ± 0.0 1.0000 8.7 ± 0.2 8.4 ± 0.3 0.2718 33.4 ± 1.1 34.2 ± 2.5 0.5436 3.8 ± 0.1 3.8 ± 0.1 0.8961 4.1 ± 0.1 3.7 ± 0.1 0.0221 68.6 ± 2.5 70.0 ± 1.5 0.4346 1.0 ± 0.0 1.1 ± 0.1 0.3539 fruto central de la segunda y última mano y número de frutos de la última mano (Tabla 1). Ambos híbridos mostraron un alto porcentaje de raíz funcional, ausencia de R. similis y bajas poblaciones de Helicotylenchus spp. y Pratylenchus spp. La población de Meloydogine fue media en FHIA-23 y baja en SH-3436-9 (Tabla 3). Ambos híbridos presentaron en promedio, un bajo coeficiente de infestación de picudos en el cormo (2.3% en FHIA-23 y 1.8% en SH-3436-9). Adicionalmente, no hubo presencia de pudriciones bacterianas asociadas. Un 66% de los cormos de FHIA-23 y un 75% de los cormos de SH-3436-9 estuvieron completamente libres de daños por picudos. Discusión Dado que el experimento consideró solo tres ciclos de cultivo, es probable que dicha cantidad de generaciones no fuera lo suficientemente amplia como para determinar diferencias entre los dos sistemas de deshije y variaciones en la composición y distribución de la población de plantas en el tiempo. No obstante, este resultado indica que hasta un tercer ciclo de cultivo, independientemente del sistema de deshije, el desempeño vegetativo y productivo y la reacción a la Sigatoka negra del hijo de sucesión en ambos híbridos es muy similar. Al igual que lo encontrado por Orellana et al. (1999), Alvarez (1977) y Orjeda (2000), FHIA-23 y SH-3436-9 presentan plantas robustas, muy altas y con un prolongado ciclo vegetativo comparado con los bananos tipo Cavendish. Lo anterior y particularmente la altura, reducen la facilidad de manejo de estos híbridos (Daniells y Bryde 1993), condición que se asocia, de acuerdo con dichos autores, a la gran pérdida de plantas por viento en la segunda generación. No obstante, Orellana et al. (1999) indican que los nuevos híbridos soportan mejor, sin necesidad de tensores, los efectos del viento. En el presente trabajo no se observó tendencia al volcamiento de estos híbridos. Sin embargo, el área de estudio no se caracteriza por presentar vientos fuertes y además las plantas fueron apuntaladas poco después de la floración. La mayor productividad de FHIA-23 estuvo fundamentalmente dada, en concordancia con Orellana et al. (1999), por una mayor cantidad de manos por racimo. La reducida cantidad de hojas a la floración en ambos híbridos se debió al efecto nocivo causado por la Sigatoka negra y a una tendencia al doblamiento prematuro de las hojas. Observaciones similares fueron hechas por Guzmán y Romero (1996) en los híbridos FHIA-01 y FHIA-02. Esta debilidad en los pecíolos, de acuerdo con Simmonds (1952) y Stover y Buddenhagen (1986), es una característica desventajosa de los tetraploides mejorados. En estudios previos desarrollados en Costa Rica por Guzmán (2000a y 2000b), ambos híbridos se mostraron medianamente resistentes a la Sigatoka negra. No obstante dada la alta severidad de la enfermedad Tabla 3. Peso de las raíces funcionales y poblaciones de nématodos (n=4) en los híbridos FHIA-23 y SH-3436-9 detercera generación. Híbrido FHIA-23 SH-3436-9 Raíz funcional por planta g % 90.0 94.0 91.0 96.8 InfoMusa - Vol. 13 - No.1 Nemátodos por 100 g de raíz Radopholus 0 0 Helicotylenchus 1800 2000 Meloidogyne 4300 100 Pratylenchus 300 0 23 que se encontró en esta investigación, es posible señalar que tanto FHIA-23 como SH-3436-9 fueron poco resistentes e iguales en su reacción al ataque del patógeno, bajo las condiciones de clima y suelo en que se desarrolló el experimento. Esta condición ya había sido anticipada por Guzmán et al. (2000), quienes encontraron, bajo condiciones de campo en parcelas de más de 100 plantas, que una alta proporción de infecciones en las hojas de estos híbridos lograban completar su ciclo y liberar inóculo secundario. Dichos autores señalaron el riesgo y la posibilidad de que la frecuencia de aislamientos capaces de romper la resistencia aumentara con el tiempo y, como consecuencia, la severidad de la enfermedad se incrementara. Lo anterior reviste particular importancia para los sistemas de manejo intensivo del cultivo (monocultivo y áreas grandes de siembra). Bajo esta perspectiva y de acuerdo a la baja cantidad de área foliar efectiva de ambos híbridos, tanto a la floración como a la cosecha, sería imprescindible considerar una estrategia de combate químico de la Sigatoka negra, aunque probablemente menos intensiva que lo comúnmente realizado en los bananos del subgrupo Cavendish, los cuales bajo estas mismas condiciones llegan a la cosecha prácticamente sin hojas (Guzmán y Romero 1998). Considerando las menores características de calidad de fruta de los híbridos tetraploides con respecto a los bananos Cavendish de exportación (Shepherd et al. 1986, Stover y Buddenhagen 1986), el potencial de ambos materiales estaría orientado dentro de un sistema de manejo para consumo local. Dada la resistencia de FHIA-23 a Fusarium oxysporum f.sp. cubense así como las bajas o ausentes poblaciones de nematodos y el bajo coeficiente de infestación de picudos encontrados en este trabajo para ambos híbridos, su cultivo para este fin mediante un uso restringido de agroquímicos sería posible, cuya factibilidad en última instancia dependería de la aceptación por parte del consumidor local. Referencias Los autores trabajan a la Dirección de Investigaciones, CORBANA S.A., Apdo 390-7210, Guápiles, Limón, Costa Rica. E-mail: [email protected]; [email protected] Alvarez J. 1997. Introducción, evaluación, multiplicación y diseminación de híbridos FHIA en Cuba. INFOMUSA 6(2):10-14. Araya M. & A. Cheves. 1998. Selección del tipo de planta para el muestreo de nematodos en banano (Musa AAA). INFOMUSA 7(1):23-26. Daniells J. & N. Bryde. 1993. Yield and plant characteristics of seven banana hybrids from Jamaica and Honduras in North Queensland. INFOMUSA 2(1):18-20. Fouré E. 1985. Black leaf streak disease of bananas and plantains (Mycosphaerella fijiensis MORELET). 24 Study of the symptoms and stages of the disease in Gabon. CIRAD-IRFA, París. 20pp. Gauhl F. 1989. Epidemiología y ecología de la Sigatoka negra (Mycosphaerella fijiensis Morelet) en plátano (Musa sp.) en Costa Rica. Tesis Ph.D. Universidad de Gottingen. Trad. Jaime Espinoza. Unión de Países Exportadores de Banano (UPEB). 128pp. Guzmán R. 2000a. Results and discussion IMTP black Sigatoka (1sT cycle) Guápiles, Costa Rica. Pp. 131149 in Evaluating bananas: a global partnership (G. Orjeda, comp.) INIBAP. Montpellier, France. Guzmán R. 2000b. Results and discussion IMTP black Sigatoka (2nd cycle) Guápiles, Costa Rica. Pp. 150163 in Evaluating bananas: a global partnership (G. Orjeda, comp.) INIBAP. Montpellier, France. Guzmán M. & R.A. Romero. 1996. Severidad de la Sigatoka negra (Mycosphaerella fijiensis Morelet) en los híbridos FHIA-01 y FHIA-02. CORBANA 21(45):41-49. Guzmán M. & R.A. Romero. 1998. Evaluación de cuatro aceites agrícolas utilizados para el combate de la Sigatoka negra (Mycosphaerella fijiensis Morelet) en el cultivo del banano (Musa AAA, cv. Gran enano). Pp. 87-89 in Informe anual 1997. Dirección de Investigaciones. CORBANA, San José, Costa Rica. Guzmán M., R. Villalta & G. Murillo. 2000. Estabilidad de la resistencia a la Sigatoka negra en híbridos de banano (Musa AAAA) con un nivel de resistencia intermedio al patógeno. Pp.31-33 in Informe anual 1999. Dirección de Investigaciones. CORBANA, San José, Costa Rica. Marín D.H, R.A. Romero, M. Guzmán & T.B. Sutton. 2003. Black Sigatoka: An increasing threat to banana cultivation. Plant Disease 87(3):208-222. Orellana P. P. Alvarado, J.Guijarro, J. Pérez, L. Pérez, P. Rowe, E. Moreno, J. Clavero, C. Romero & M. Hernández.. 1999. Introducción y validación de híbridos tetraploides de Musa en Cuba. CORBANA 24(51):79-84 Orjeda G. 2000. Evaluating bananas: a global partnership. Results of IMTP phase II. INIBAP. Montpellier, France. 466pp. Orjeda G., J.V. Escalant & N. Moore. 1999. Programa Internacional de Evaluación de Musa (IMTP) fase II - sinopsis del informe final y resumen de los resultados. INFOMUSA 8(1):3-10. Pérez L. 2000. Prácticas culturales para aumentar la productividad. Deshija unidireccional. Pp. 6-7 in Informe Anual 1999. Dirección de Investigaciones. CORBANA, San José, Costa Rica. Rivera M. & J. Dueñas. 2002. Reacción a pudrición de corona de frutos del banano híbrido FHIA-23. Pp. 29-32 in Programa de Banano y Plátano. Informe Técnico 2001. Fundación Hondureña de Investigación Agrícola (FHIA), Honduras. 35pp. Romero R. 1994. Calculation of infection index. Technical Guidelines for IMTP Phase II: Sigatoka Sites. Pp. 277 in The improvement and testing of Musa: a global partnership (D.R. Jones, ed.). Proceedings of the First Global Conference of the International Musa Testing Programme held at FHIA, Honduras. INIBAP, France. Rowe P. 1998. Breeding bananas and plantain resistant to diseases and pest. Pp. 54-60 in Organic/environmentaly friendly banana production. Proceedings of a workshop held at EARTH (F. Rosales, S. Tripon & J. Cerna, eds). INIBAP;IDRC; EARTH. Guácimo, Costa Rica. 250pp. Rowe P. 1999. Mejoramiento de bananos y plátanos para resistencia a enfermedades: eventos cruciales y sus implicaciones. CORBANA 24(51):99-110. InfoMusa - Vol. 13 - No.1 Shepherd K., J.L. Loyola & E.J. Alves. 1986. Mejoramiento genético del banano. Pp. 1-19 in Mejoramiento genético de banano y plátano en Brasil y Honduras. Unión de Países Exportadores de Banano. Panamá. Simmonds N. 1952. The strength of banana petiols in relation to ploidy. Annals of Botany XVI(63): 341-347. Stover H. & I. Buddenhagen. 1986. Fitomejoramiento del banano: poliploidia, resistencia a enfermedades y productividad. Pp. 20-55 in Mejoramiento genético de banano y plátano en Brasil y Honduras. Unión de Países Exportadores de Banano. Panamá. Stover R.H. & N.W. Simmonds. 1987. Bananas. 3rd ed. Longman Scientific and Technical. New York. 468pp. Vilardebo A. 1973. Le coefficient d’infestation, critère d’evaluation du degré d’attaques des bananeraies par Cosmopolites sordidus GERM. le charançon noir du bananier. Fruits 28(6):417-426. Evaluación de nuevos híbridos de banano contra la enfermedad de la Sigatoka negra Híbridos mejorados V. Krishnamoorthy, N. Kumar, K. Angappan y K. Soorianathasundaram L a enfermedad de la raya negra de la hoja o Sigatoka negra, causada por Mycosphaerella fijiensis Morelet, es una de las enfermedades fungosas más serias que afecta la producción de banano y plátano (Stover y Buddenhagen 1986). El desarrollo de variedades resistentes se considera como la tecnología más apropiada para controlar esta enfermedad de la mancha foliar. La Universidad Agropecuaria de Tamil Nadu inició un programa de mejoramiento genético para incorporar resistencia duradera a la Sigatoka negra en bananos, utilizando bananos diploides altamente resistentes e híbridos sintéticos. El estudio actual se realizó para evaluar las respuestas de los nuevos híbridos a la enfermedad de Sigatoka negra. Materiales y métodos El estudio fue conducido en el Departamento de cultivos frutícolas de la Universidad Agropecuaria de Tamil Nadu en Coimbatore, India, entre junio de 2000 y febrero de 2002. Coimbatore esta localizado a 11oN y 77oE y a una altitud de 427 msnm. Las temperaturas máxima y mínima son 31°C y 21°C, respectivamente, la humedad relativa es 85% en la mañana y 49% en la tarde, y la precipitación anual durante el experimento fue de 698 mm. La textura del suelo es franco arenosa arcillosa. Los híbridos evaluados en este estudio se presentan en la Tabla 1. Estos híbridos fueron cribados bajo condiciones naturales de campo y sembrados en el campo como se describe en Orjeda et al. (1998). Se seleccionaron retoños con tamaño uniforme, los cuales fueron sembrados en hoyos de un pie cúbico con un espaciado de 1.8 m x 1.8 m. Las plantas fueron regadas diariamente con 20 litros de agua a través de un sistema de goteo y se aplicaron InfoMusa - Vol. 13 - No.1 110:35:330 g de N:P:K. El nitrógeno y potasio fueron aplicados a través de un sistema de goteo con una unidad de fertigación a intervalos semanales en 36 raciones iguales empezando desde la 9a semana. No se aplicaron medidas de control químico, biológico o cultural contra la Sigatoka negra. El deshije se realizó a intervalos mensuales para fomentar el crecimiento de la planta madre. El diseño experimental fue un diseño de bloques completamente al azar con cinco réplicas para cada híbrido y clon parental. La severidad de la enfermedad o la cantidad de área foliar afectada fue expresada como grados (de 0 a 6) utilizando la modificación de Gauhl de la escala de Stover (Gauhl 1994). Se registraron los siguientes parámetros: la cantidad de hojas funcionales y la hoja más joven manchada, es decir el numero de hoja de la hoja más joven manchada con 10 o más lesiones maduras (Vakili 1968). I. I. = Σnb (N –1)T Tabla 1. Parentesco, ploidia y genoma de los híbridos de banano evaluados. Número del híbrido H-59 H-65 H-66 H-89 H-110 H-201 H-203 H-204 H-205 H-208 H-209 H-210 H-211 H-02-01 H-02-08 H-02-11 H-02-12 Parentesco Matti (AA) × Anaikomban (AA) Matti (AA) × Pisang lilin (AA) Matti (AA) × Anaikomban (AA) Matti (AA) × Namarai (AA) Matti (AA) × Tongat (AA) Bareli chinia (ABB) × Pisang lilin (AA) × Robusta (AAA) H-59 (AA) × Ambalakadali (AA) H-65 (AA) × Pisang lilin (AA) H-66 (AAA) × Ambalakadali (AA) H-89 (AA) × Anaikomban (AA) H-201(AB) × Ambalakadali (AA) H-201 (AB) × Anaikomban (AA) H-201 (AB) × Pisang lilin (AA) Ambalakadali (AA) × Anaikomban (AA) H 201 (AB) × Eraichivazhai (AA) H-201 (AB) × H-110 (AA) H-201 (AB) × H-110 (AA) 25 Ploidia 2× 2× 3× 2× 2× 2× 2× 2× 2× 2× 3× 3× 2× 2× 2× 2× 2× Genoma AA AA AAA AA AA AB AA AA AA AA AAB AAB AA AA AB AB AB El índice de infección (I.I.) fue calculado como sigue: I. I. = donde n: número de hojas en cada grado b: grado N: número de grados utilizados en la escala T: total número de hojas anotada La resistencia fue evaluada midiendo el desarrollo de la enfermedad (en este caso el índice de infección). La resistencia total (inmunidad) ocurría cuando la enfermedad no fue capaz (índice de infección = 0) de desarrollarse en el tejido de la planta. La resistencia parcial ocurría cuando el desarrollo de la infección y de la enfermedad era limitado, en comparación con las plantas susceptibles. El índice de la hoja más joven manchada fue calculado como sigue: IHMJM = T- HMJM -1 x 100 T donde HMJM: hoja más joven manchada T: número total de hojas. El índice de área foliar no manchada fue calculado como sigue: INHE = HMJM -1 x 100 NHE donde NHE: número de hojas erectas. Tabla 2. Desempeño agronómico durante le floración de los híbridos y clones parentales expuestos a Mycosphaerella fijiensis (n=5). Número de hojas por planta Hoja más joven manchada Indice de Indice de Indice de infección hoja más joven área foliar manchada no manchada Híbridos H-203 13.2 bc H-204 17.1 ab H-205 13.2 bc H-208 17.2 ab H-209 19.3 a H-210 15.1 b H-211 18.2 ab H-02-01 14.1 bc H-02-08 17.3 ab H-02-11 15.4 b H-02-12 11.5 cd Clones parentales Ambalakadali 15.3 b Anaikomban 14.2 bc Eraichivazhai 15.7 b Pisang lilin 10.2 d H-59 13.3 bc H-65 11.3 cd H-66 13.7 bc H-89 9.8 d H-110 13.5 bc H-201 13.6 bc Rasthali (testigo) 12.0 c 11.1 cd 16.2 b 12.1 bc 14.2 b 19.3 a 15.3 b 14.4 b 6.7 e 16.2 b 12.1 bc 11.2 cd 3.8 b 2.0 b 2.6 b 4.9 b 0.9 a 1.1 a 5.6 b 23.8 d 2.0 b 4.4 b 1.5 b 23.1 c 11.8 de 15.4 d 23.5 c 5.3 e 6.7 e 27.8 c 64.3 b 11.7 de 26.7 c 9.1 de 76.9 ab Tolerante 88.2 ab Tolerante 84.6 ab Tolerante 76.5 ab Tolerante 94.7 a Resistant 93.3 a Resistant 72.2 ab Tolerante 35.7 d Altamente suceptible 88.2 ab Tolerante 73.3 ab Tolerante 90.9 a Tolerante 11.2 cd 11.2 cd 13.1 bc 10.3 cd 12.0 bc 10.0 cd 13.2 bc 8.3 d 13.4 bc 12.5 bc 3.3 f 8.9 c 8.3 c 4.4 b 1.7 b 2.6 b 3.0 b 1.3 a 3.7 b 1.3 a 2.6 b 74.0 e 33.3 c 28.6 c 20.0 cd 10.0 de 15.4 d 18.2 cd 7.7 e 22.8 c 7.7 e 15.4 d 80.3 a 66.7 bc Susceptible 71.4 ab Susceptible 80.0 ab Tolerante 90.0 a Tolerante 84.6 ab Tolerante 81.8 ab Tolerante 92.3 a Resistant 77.8 ab Tolerante 92.3 a Resistant 84.6 ab Tolerante 19.2 e Altamente suceptible Los promedios seguidos por la misma letra no difieren significativamente (p≤0.05) de acuerdo con la prueba HSD de Tukey. 26 El índice de infección, el índice de la hoja más joven manchada y el índice del área foliar no manchada fueron transformados mediante el arcoseno y los datos fueron sujetos a un análisis de varianza. Los promedios se compararon utilizando la prueba de proporción K de Waller Duncan a p<0.05. En las hojas en el período de floración se midieron varios componentes bioquímicos y actividades enzimáticas utilizando los métodos descritos en la literatura: contenido clorofílico (Yoshida et al. 1971), proteína soluble total (Lowry et al. 1951), pralina (Bates et al. 1973), fenoles totales (Spies 1957), fenol OD (Arnow 1937), fenol ligado (Chattopadhyay y Samadar, 1980), ácido clorogénico y taninos (Sadasivam y Manickam 1997), lignina (Chesson 1978), cera epicuticular (Ebercon et al. 1977), actividad de la peroxidasa (Hartec 1955), actividad de la polifenol oxidasa (Mayer et al. 1965), actividad de la fenilalanina amoníaco liasa (Ross y Sederoff 1992), actividad de la catalasa (Luck 1974) y la actividad de la oxidasa de ácido ascórbico (Sadasivam y Manickam 1997). Resultados y discusión En los bananos, el desarrollo de los racimos depende del potencial fotosintético de las hojas. Las plantas de banano requieren más de 70% de follaje activo y un mínimo de 8 hojas funcionales para el desarrollo adecuado de las frutas (Orjeda 1998). En el presente estudio, no se observó resistencia total en los híbridos y cultivares parentales investigados (Tabla 2). Se descubrió que H-209 y H-210 y los clones parentales H-66 y H-110 fueron resistentes a la Sigatoka negra como resultado de su bajo índice de infección y alto índice de área foliar no manchada. ‘Anaikomban’ y ‘Ambalakadali’ fueron clasificados como susceptibles debido a su alto índice de infección y valor relativamente bajo de la hoja más joven manchada, mientras que H-02-01 fue clasificado como altamente susceptible debido a su índice de infección muy alto y pequeña proporción de área foliar libre de rayas y manchas. Se estudió la relación entre el índice de infección y varias sustancias bioquímicas. Los niveles de clorofila, azúcares reductores y azúcares totales, y la actividad de la oxidasa de ácido ascórbico estaban positivamente correlacionados con el índice de infección, mientras que los niveles de prolina y lignina, y la actividad de la peroxidasa estaban negativamente correlacionados con el índice de infección (Tabla 3). Los niveles de los azúcares reductores, azúcar total y lignina, y InfoMusa - Vol. 13 - No.1 la actividad de la fenilalanina amoníaco liasa, oxidasa de ácido ascórbico y catalasa estaban negativamente correlacionados con el valor para la hoja más joven manchada. El área foliar sin rayas foliares estaba positivamente correlacionada con las concentraciones de los azúcares totales, cera, fenoles y lignina, así como con la actividad de la fenilalanina amoníaco liasa, oxidasa de ácido ascórbico, catalasa y fenol oxidasa. Referencias Arnow L.E. 1937. Colorimetric determination of the components 2,4 - dihydroxy phenol alanine and tyrosine mixtures. J. Biol. Chem. 18:537. Bates L.S., R.P. Waldren & T.D. Teare. 1973. Rapid determination of free proline for water stress studies. Plant and Soil 39:205-207. Chattopadhyay A.D. & K.R. Samadar. 1980. Effects of Helminthosporium oryzae infection and ophiobilin on the phenol metabolism of host tissues. Phytopathol. 98:193-202. Chesson A. 1978. The maceration of line flax under anaerobic condition. J. Appli. Bacteriol. 45:219-230. Ebercon A., A. Blum & W.R. Jordan. 1977. A rapid colorimetric method for wax content of sorghum leaves. Crop Sci. 17:179-180. Gauhl F. 1994. Epidemiology and ecology of black Sigatoka (Mycosphaerella fijiensis Morelet) on plantain and banana in Costa Rica. INIBAP, Montpellier, France. 120pp. Hartec E.F. 1955. Haematin compounds. Pp. 197-245 in Modern Methods of plant analysis Vol.4 (K.Peach & M. Tracq, eds). Springer-Verlag, New York. Lowry O.H., J. Rosebrough, L.A. Farr & R.J. Randael. 1951. Protein measurement with folin phenol reagent. J. Biol. Chemi. 193:265-275. Luck H. 1974. Methods in enzymatic analysis. Academic Press, New York, p. 885. Mayer A.M., E. Haul & R.B. Shaul. 1965. Assay of catechol oxidase, a critical comparison of method. Phytochem. 5:783-789. Orjeda G. 1998. Evaluación de la resistenciaa de los bananos a las enfermedades de Sigatoka y marchitamiento for Fusarium. Guías técnicas INIBAP 3. INIBAP, Montpellier, France. Ross W.W. & R.R. Sederoff. 1992. Phenylalanine ammonia lyase from lobololly pine. Purification of the enzyme and isolation of complementary DNA clones. Plant Physiol. 98:380-386. Sadasivam S. & A. Manickam. 1997. Biochemical methods. New Age International Publishers, New Delhi. Spies J.R. 1957. Colorometric procedure for amino acids. Methods in Enzymology (3): 467-477. Stover R.H. & I.W.Buddenhagen. 1986. Banana breeding, polyploidy disease resistance and productivity. Fruits 41:175-191. Vakili N.G. 1968. Response of Musa acuminata species and edible cultivars to infection by Mycosphaerella musicola. Trop. Agr. (Trinidad) 45:13.22. Tabla 3. Coeficientes de correlación de los parámetros de la enfermedad con los niveles de las sustancias bioquímicas y la actividad enzimática. Parámetros Clorofila total Proteína soluble total Azúcares reductores Azúcares totales Prolina Cera Fenol total Fenol OD Fenol ligado Lignina Fenilalanina amoníaco liasa Oxidasa de ácido ascórbico Catalasa Polifenol oxidasa Peroxidasa Indice de infección 0.540** 0.311 0.494** 0.550** -0.444* 0.138 0.260 -0.142 0.269 -0.445* 0.282 0.515** 0.332 0.327 -0.431* Hoja más joven manchada 0.020 -0.012 -0.442* -0.515** 0.480* -0.476* -0.384 -0.375 -0.445* -0.519** -0.524** -0.549** -0.524** -0.362 0.663** S. Coulibaly y C. Djédji E InfoMusa - Vol. 13 - No.1 Indice de hoja más joven manchada -0.068 -0.053 -0.481* -0.594** 0.542** -0.548** -0.439* -0.384* -0.560** -0.592** -0.562** -0.612** -0.581** -0.438* 0.746** Indice de area foliar no manchada 0.068 0.052 0.481* 0.593** -0.540** 0.548** 0.442* 0.386* 0.562** 0.592** 0.564** 0.606** 0.582** 0.440* -0.748** *Significativo a probabilidad de 0.05 ** Significativo a probabilidad de 0.01 Cualidades organolépticas de los frutos de los híbridos SH-3640 y CRBP-39 n Côte d’Ivoire, el programa de investigación de frutos y cítricos del Centre national de recherche agronomique (CNRA) lleva a cabo actualmente un estudio sobre el comportamiento varietal del híbrido SH-3640 (AAAA), banano de postre procedente del programa de creación varietal de la Fundación Hondureña de Investigación Agrícola (FHIA) en Honduras, y del CRBP-39 (AAAB), plátano híbrido procedente del programa de creación varietal del Centre Africain de Recherches sur Bananiers et Plantains (CARBAP) en Camerún. Los autores trabajan en el Department of Fruit Crops, Horticultural College and Research Institute, Tamil Nadu Agricultural University, Coimbatore – 641003, India. La dirección de V. Krishnamoorthy es Krishi Vigyan Kendra, Vriddhachalam, Cuddlore, Tamil Nadu, India. Estos híbridos mostraron una resistencia parcial a la Sigatoka negra. Complementando dicho estudio, el centro de investigación tecnológica del CNRA dirigió un estudio sobre la evaluación sensorial de estos híbridos para determinar su aceptabilidad por los consumidores marfileños. La evaluación sensorial de nuevos híbridos y de nuevos cultivares es una etapa decisiva en el proceso de selección y creación varietal. Esta etapa, situada entre la producción de los bananos en la estación experimental y la difusión en el medio campesino de las nuevas plántulas 27 Evaluación sensorial ermite pronunciarse sobre la aceptabilidad de nuevos productos por parte de los futuros consumidores. Material y métodos Las pruebas se efectuaron en los híbridos SH3640 y CRBP-39 puestos a nuestra disposición por el centro de investigación de frutos y cítricos del CNRA de Anguédédou, cerca de Abidjan. Se cosecharon 338 días después de establecer el ensayo y en su estado de máxima maduración, es decir, a partir de la aparición de un dedo amarillo en el racimo (Mitra 1997). Como muestras de referencia (Watts et al. 1991), se emplearon bananos de postre (‘Poyo’, AAA) y plátanos (de tipo False horn, AAB) comprados en el mercado y con el mismo grado de madurez que los híbridos con los que se compararon (IPGRI-INIBAP/CIRAD 1996). Para la comparación de los bananos de postre, los bananos se pelaron a mano y se cortaron en rodajas de 1 cm de grosor con un cuchillo de acero inoxidable. Todas las pruebas de evaluación se efectuaron en los 10 minutos que siguieron al corte. Una evaluación preliminar hecha por dos investigadores del centro había permitido determinar las distintas preparaciones culinarias posibles: el foutou o plátano machacado, el foufou o plátano triturado, el aloco o plátano frito, y los chips. Estos platos se hacen del siguiente modo: El foutou Los dedos de banano se lavan, se pelan y se parten en dos, con un cuchillo de acero inoxidable, para extraer los puntos negros (semillas atrofiadas). Seguidamente, se cortan en rodajas homogéneas de 3 cm de grosor y se cuecen 20 mn en agua hirviendo. Una vez frías, se machacan las rodajas en un mortero tradicional de madera hasta obtener una pasta blanda de consistencia pegajosa, llamada foutou (Mosso et al. 1996). El foufou El método de preparación es idéntico al del foutou con la diferencia de que, en vez de machacarse, las rodajas cocidas se desmenuzan en el mortero y se mezclan luego con aceite de palma (aprox. 0,25 L para 1 kg de banano cocido). Se obtiene así un puré llamado foufou. El aloco Tras su lavado y pelado como en los casos anteriores, la pulpa del banano se corta en rodajas homogéneas de 1 cm de grosor. A continuación, se fríen las rodajas en aceite vegetal a 160-180°C durante 5 minutos (Tchango Tchango y Ngalani 1998). Las rodajas deben quedar blanditas. 28 Los chips Para preparar los chips, se procede de la misma manera que para el aloco. La diferencia estriba en que las rodajas son más finas (unos 0.3 cm solamente) y en que se fríen durante menos tiempo (2 a 3 minutos) (Tchango Tchango y Ngalani 1998). Los chips deben quedar crujientes. Se compararon los chips preparados a partir del híbrido SH-3640, en estado de maduración “amarillo”, y del híbrido CRBP-39, en estado de maduración “verde claro”, con chips hechos con plátanos de tipo False horn en los mismos estados de maduración y comprados en el mercado. En lo que respecta a otros métodos de cocción, solamente se evaluaron las calidades culinarias del híbrido CRBP-39. El panel de degustadores Para platos como los chips y el aloco, de fácil preparación, se formó un panel de degustadores a partir de una selección de empleados del CNRA de Abidjan, compuesta por una decena de personas de ambos sexos, de distintas clases sociales y con una edad comprendida entre 35 y 50 años. Éstos respondieron a un cuestionario adaptado a las particularidades de los platos que debían evaluarse. La mitad de los panelistas ya había participado en pruebas de degustación. Para los demás platos (foutou y foufou), se eligieron cinco hogares. La elección de los hogares se efectuó según los siguientes criterios: 1°) consumir plátanos frecuente y regularmente, 2°) poseer un nivel y una calidad de vida correctos para la Côte d’Ivoire, 3°) no superar las cinco personas en el hogar. Los hogares realizaron sus comentarios por escrito, una vez informados de los criterios que debían tenerse en cuenta, en particular, la elasticidad, el carácter pegajoso, la apetencia y la firmeza de la pasta. Las pruebas de evaluación Las muestras, identificadas por un código de tres cifras, fueron presentadas en orden aleatorio a los degustadores (Watts et al. 1991). Se trataba de pruebas de medidas de intensidad de seis niveles, en las que todas las muestras se presentaron al mismo tiempo a los degustadores para poder evaluar de nuevo algunas, si fuera necesario. Las características escogidas fueron: el gusto, el color, la consistencia, la textura, el aroma y el regusto. Las notas que debían asignarse iban de 1 a 5 y correspondían a las siguientes valoraciones: 1=malo; 2=mediocre; 3=medio; 4=bueno; 5=muy bueno. La única excepción fue la evaluación del regusto, en donde una nota baja era positiva porque significaba poco InfoMusa - Vol. 13 - No.1 Tabla 1. Promedio y desviación típica de las notas (de 1 a 5) asignadas por un panel de degustadores al banano híbrido SH-3640 y Poyo, un banano de postre comprado en el mercado. SH-3640 Poyo Gusto 3.1±0.9 3.0±0.8 Color 3.2±0.9 3.0±0.7 Consistencia 3.4±0.7 3.1±0.8 regusto. En las demás características, cuanto más alta era la nota elevada, más se valoraba el banano. Se realizó un análisis de varianza (ANOVA) para determinar si existían diferencias significativas entre los tipos de bananos evaluados (Watts et al. 1991). Resultados y discusión Comparación entre bananos de postre Los valores promedio de las notas asignadas por el panel de degustación estaban muy próximos entre ellos (Tabla 1). Según estos resultados, y en los criterios de gusto, color, consistencia y textura, se prefirió el banano híbrido SH-3640 al banano de postre ‘Poyo’ comprado en el mercado. Además de su aroma, considerado inferior al de ‘Poyo’, en SH-3640 se valoró negativamente un regusto ligeramente más acentuado. Es posible que las notas correspondientes al regusto, con una escala inversa a las demás características, no fueran bien comprendidas. Del análisis de varianza (Tabla 2) de los resultados de esta prueba se desprende que, en el gusto y la consistencia, las diferencias en las calificaciones no son significativas al nivel del 5%, tanto para los tratamientos como para los degustadores. Los coeficientes F calculados son, en efecto, inferiores a los valores críticos de Snedecor. En color, textura y regusto no aparece ninguna diferencia significativa en los tratamientos. Por el contrario, se observan diferencias en los degustadores, pero siguen siendo despreciables, ya que las desviaciones entre los valores calculados de los coeficientes F y las de la tabla de Snedecor son muy bajas. Sólo el aroma, con una diferencia de 3 puntos entre los valores de F, presenta una ligera Textura 2.9±1.0 2.7±0.7 Aroma 2.6±0.8 3.0±0.9 Regusto 3.3±0.9 3.1±1.0 Total 15.2±5.2 14.8±5.9 diferencia significativa para los tratamientos al nivel del 5%. Comparación de chips El gusto y el aroma de chips de plátanos de tipo False horn comprados en el mercado en estado de maduración “amarillo” se consideraron superiores a los de los híbridos y obtuvieron notas altas (Tabla 3). Los chips preparados a partir de bananos híbridos de CRBP-39 (estado verde) fueron los menos apreciados. Los resultados del análisis de varianza muestran que las diferencias registradas en el color, la textura y el regusto no son significativas al nivel del 5% (Tabla 4). Se observan, por el contrario, diferencias significativas al nivel del 5% en el gusto, la consistencia y el aroma. Por otra parte, ninguna diferencia significativa al nivel del 5% puede atribuirse a los degustadores. Las distintas valoraciones de los hogares concuerdan completamente. Se pueden resumir del siguiente modo: Tabla 2. Análisis de varianza aplicado a los datos del panel de degustación que evalúa los bananos de postre. Característica Gusto Fuente de variación Coeficiente F Calculado 0.1020 1.9184 1.0000 5.4500 0.2301 3.4400 0.6286 3.6000 8.2051 3.6461 0.4739 3.4745 Genotipo Degustadores Genotipo Degustadores Genotipo Degustadores Genotipo Degustadores Genotipo Degustadores Genotipo Degustadores Color Consistencia Textura Aroma Regusto *Significativo a P=0.05 Tabla 3. Promedio y desviación típica de las notas (de 1 a 5) asignadas por un panel de degustadores a chips elaborados a partir de plátanos de tipo False horn e híbridos de plátano SH-3640 y CRBP-39 con diferentes estados de maduración. Chips de plátano amarillo del mercado Chips de plátano verde del mercado Chips de banano híbrido SH-3640 amarillo Chips de banano híbrido CRBP-39 verde Gusto 4.0±0.7 Color 3.8±0.6 Consistencia 3.3±1.0 Textura 3.2±1.0 Aroma 3.7±0.7 Regusto 3.8±0.4 Total 18.0±4.4 2.8±1.0 3.1±1.0 3.3±1.0 3.2±1.0 2.6±1.0 2.9±0.8 15.0±5.8 3.2±1.3 2.8±0.9 2.2±0.8 2.3±1.1 3.2±0.9 2.9±1.1 13.7±6.1 2.6±1.2 3.1±1.0 3.0±1.0 3.0±1.0 2.7±1.0 2.9±0.8 14.4±6.0 InfoMusa - Vol. 13 - No.1 29 Tabla* 5.1174 3.1789 5.5914 3.7470 5.3177 3.4381 5.3177 3.4381 5.3177 3.4381 5.1177 3.1789 Tabla 4. Resultados del análisis de varianza realizado sobre los datos del panel de degustación que evalúa los chips. Característica Gusto Color Consistencia Textura Aroma Regusto Fuente de variación Genotipo Degustadores Genotipo Degustadores Genotipo Degustadores Genotipo Degustadores Genotipo Degustadores Genotipo Degustadores Coeficiente F Calculado 3.2343 0.5250 2.3033 1.4075 4.0062 1.7076 1.7189 1.4703 5.2623 6.1934 3.3744 2.2654 Tabla* 2.9604 2.2501 2.9604 2.2501 2.9604 2.2501 3.0088 2.3551 3.0725 2.4876 3.0088 2.3551 * Significativo a P=0.05 Evaluación del híbrido CRBP-39 después de hervirlo en agua. Souleymane Coulibaly* y Catherine Djédji trabajan en la Station de Recherche Technologique (SRT) del Centre National de Recherche Agronomique (CNRA), 08 BP 881 Abidjan 08 Côte d’Ivoire. Correspondencia: Coulibaly Souleymane. E-mail: [email protected] • La maduración de los dedos en el racimo de bananos se efectúa de manera progresiva. Por ejemplo, en dos dedos contiguos de la misma hilera de una misma mano, uno puede estar en el estado de maduración “verde claro” y otro en el “amarillo”. Ahora bien, entre estos dos estados existen el “verde agua” y “verde amarillo”, según la tabla de colores de los “Descriptores para el banano” (IPGRI-INIBAP/ CIRAD 1996). También se observó un estado “verde medio” que precedía al “verde claro”. • El pelado de los frutos es fácil pero éstos no tienen mucha pulpa, en comparación con los otros cultivares. • La parte central no contiene o tiene muy pocos puntos negros (semillas atrofiadas), lo que es una ventaja considerable durante el pelado, ya que la extracción de esta parte central es indispensable para la obtención de bolitas homogéneas de foutou que tengan buen aspecto. En lo relativo a las cualidades culinarias, los hogares señalaron, tras cocción de dedos en distintos estados de maduración en agua durante 20 minutos y en el mismo recipiente, que: • los dedos en estado de maduración “amarillo” están muy blandos y son fáciles de machacar. Desgraciadamente, la pasta se pega al mortero lo que complica la elaboración de las bolitas, • los dedos en estados de maduración “verde medio” y “verde claro” son difíciles de machacar porque están muy duros, • los dedos en estado de maduración “verde amarillo” son de fácil machacado y la pasta puede trabajarse fácilmente para elaborar las bolas de foutou. Los hogares otorgaron una calificación muy buena al gusto del CRBP-39, tanto hervido como en foutou. En conclusión, estos dos híbridos presentan cualidades organolépticas indudables: el SH- 30 3640 como banano de postre y el CRBP-39 como plátano de cocción. El híbrido CRBP39 no fue apreciado en forma de chips pero, tras hervirlo en agua, se presta bien para la preparación del foutou, sobre todo cuando está en estado de maduración “verde amarillo”. Este híbrido tiene la ventaja de la maduración progresiva de los dedos de un mismo racimo. Esto hace que se puedan consumir estos bananos durante un período de tiempo más largo sin pérdidas de importancia. La continuación de estos trabajos nos permitirá determinar sus cualidades nutritivas. Agradecimientos Queremos expresar nuestro sincero agradecimiento al Dr Kobena Kouman, investigador en la Estación de Bimbresso del CNRA, por su decisiva contribución en el marco de este estudio. Referencias IPGRI – INIBAP/CIRAD. 1996. Descriptores para el Banano (Musa spp.). IPGRI, Roma, Italia; INIBAP, Montpellier, Francia; CIRAD, Montpellier, Francia. Mitra G.S.K. (ed.). 1997. Postharvest physiology and storage of tropical and subtropical fruits. CAB International, Wallingford, United Kingdom. 423pp. Mosso K., N. Kouadio & G.J. Nemlin. 1996. Transformation traditionnelle de la banane, du manioc, du taro et de l’igname dans les régions du centre et du sud de la Côte d’Ivoire. IAA (mars): 91-96. Tchango Tchango J. & J.A. Ngalani. 1998. Transformations et utilisations alimentaires de la banane plantain en Afrique centrale et occidentale. Pp. 361-373 in Bananas and food security - Les productions bananières : un enjeu économique majeur pour la sécurité alimentaire. International symposium, Douala, Cameroun, 10 – 14 novembre 1998. (C. Picq, E. Fouré and E. A. Frison, eds). INIBAP, Montpellier, France. Watts B.M., G. L. Ylimaki, L. E. Jeffery & L. G. Elias. 1991. Méthodes de base pour l’évaluation sensorielle des aliments. CRDI, Ottawa, Canada. InfoMusa - Vol. 13 - No.1 Análisis preliminar de la literatura científica sobre las especies silvestres de Musa Se analizó la información contenida en una selección de artículos científicos dedicados a la descripción de bananos silvestres y a su distribución para desarrollar una base de datos bibliográficos sobre los bananos silvestres. Se consultaron diferentes bases de datos (MusaLit, CAB-Abstract, Current Contents, etc.) para elaborar esta lista. Algunos expertos en taxonomía del banano, especialmente el Dr Edmond de Langhe y el Sr Markkü Häkkinen, aportaron la información complementaria que habían reunido en las distintas misiones efectuadas en Indonesia y Tailandia. Esta acción tenía como objetivo final la integración de dicha información en el Sitema de información sobre el germoplasma de Musa (Musa Germplasm Información System, MGIS), una base de datos desarrollada y administrada por INIBAP. Esta base de datos contiene las descripciones de más de 5000 accesiones conservadas en bancos de genes de todo el mundo http://mgis.grinfo.net) con una presencia mayoritaria de variedades cultivadas localmente. Las especies silvestres se encuentran subrepresentadas, de ahí la elaboración de este proyecto destinado a analizar documentos científicos referentes a las especies silvestres reportadas u observadas por botánicos en misiones de recolección o encuestas con el fin de, entre otras cosas, determinar su distribución. No obstante, la información geográfica contenida en estos documentos suele ser imprecisa. Por lo tanto, sólo se añadieron unos 15 artículos Focus sobre los bananos silvestres al MGIS. Este estudio preliminar deberá completarse para ampliar el ámbito de literatura científica que sirve de base a la integración de datos publicados en la base de datos. Dado que el MGIS fue concebido para incorporar datos científicos relativos al material vegetal conservado en colecciones, y no los tomados de la literatura, se elaboró una lista de recomendaciones para que esta herramienta pudiera admitir la información procedente de la literatura científica. Los datos incorporados al MGIS (coordenadas geográficas, información sobre las poblaciones de bananos, etc.) se transfirieron seguidamente al software de información geográfica DIVA-GIS. Dicho programa, concebido por Robert Hijmans en el marco de su trabajo en el CIP (Centro Internacional de la Papa), es de distribución gratuita (http://www.diva-gis.org/) y permite la elaboración de mapas geográficos centrados especialmente en la biodiversidad. Se elaboraron mapas de la distribución y la diversidad genética del banano en distintos países de Asia y del Pacífico. Este trabajo permitió mejorar nuestros conocimientos, puesto que el último mapa que reflejaba la distribución del género Musa, fechado en 1967, se lo debemos a Jean Champion (Figura 1). Desde entonces, se han observado especies pertenecientes a la sección Callimusa en China (Liu et al. 2002) y especies pertenecientes a la sección Australimusa en Borneo (Hotta 1967). Este trabajo nos permitió comprobar que algunos países como Birmania, Camboya y Figura 1. Distribución de las cuatro secciones de Musaceae según Champion 1967 (adaptado por Guinard 2002). InfoMusa - Vol. 13 - No.1 31 Patrick Pollefeys realizó este trabajo en el ámbito del programa de prácticas de seis meses en organizaciones internacionales gubernamentales financiado por el Ministerio de Relaciones Internacionales de Quebec. El trabajo fue dirigido por Suzanne Sharrock, ex responsable del programa de conservación de la diversidad de los bananos en el INIBAP y Elizabeth Arnaud, responsable del MGIS. Los resultados de este estudio y la bibliografía utilizada se presentan en el informe «Preliminary analysis of the literature on the wild Musa species distribution using MGIS and DIVA-GIS» disponible en sitio web del INIBAP en http: //www.inibap.org/publications/ reports_eng.htm Focus sobre banano orgánico Laos – a pesar de encontrarse en el centro de diversidad del género Musa - poseen escasos datos sobre sus especies de bananos indígenas. Además, parece que el hábitat de algunas especies se reduce con el paso del tiempo. Por ejemplo, una misión dirigida por Argent (1976) a mediados de los setenta en Papúa Nueva Guinea reveló la presencia de Musa balbisiana en la provincia de Morobe; Sharrock (1989), al frente de otra misión de recolección en 1988-89, no detectó su presencia. Durante esta misión, sólo se pudo muestrear Musa balbisiana en la isla de Nueva Bretaña, lo que indica una reducción de su distribución con relación a los años anteriores. La presencia de Musa coccinea en China se vuelve cada vez más escasa debido a la actividad humana. Esta especie se distribuía ampliamente en los bosques de la provincia de Yunnan pero, en una reciente misión de recolección, los científicos chinos tuvieron dificultades para localizarla (Pollefeys et al. 2004). La elaboración de mapas con el programa DIVA permitió también visualizar el esquema de distribución de la sección Australimusa en Papuá Nueva Guinea. Por ejemplo, se observa que Musa boman y Musa lolodensis se encuentran en la parte noroeste de la isla, que Musa peekelii ssp. angustigemma se limita a la provincia de Madang y de Eastern Highlands, mientras que M. peekelii ssp. peekelii parece hallarse sólo en la isla de Nueva Irlanda (Argent 1976, Sharrock 1989). Este tipo de información puede servir de guía a futuras misiones de recolección para orientar sus investigaciones hacia regiones en las que la distribución de Musa es menos conocida y, eventualmente, para desarrollar estrategias de conservación en su hábitat natural. Referencias Argent G.C.G. 1976. The wild bananas of Papua New Guinea. Notes from the Royal Botanic Garden Edinburgh 35:77-114. Champion J. 1967. Les bananiers et leurs cultures. Edition Setco, Paris. Guinard O., E. Arnaud & S. Sharrock. 2002. Preliminary analysis of Musa Germplasm Information System data for Southeast Asia using the Geographical Information System Software DIVA-GIS: a data analysis. INIBAP, Montpellier, France. Hotta M. 1967. Notes on the wild bananas of Borneo. Journal of Japanese Botany 42:344-353. Liu A.Z., D.Z. Li and X.W. Li. 2002. Taxonomic notes on wild bananas (Musa) from China. Botanical Bulletin of Academia Sinica 43:77-81. Pollefeys P., S. Sharrock & E. Arnaud. 2004. Preliminary analysis of the literature on the distribution of wild Musa species using MGIS and DIVA-GIS. INIBAP, Montpellier, France. Sharrock S. 1989. Report on the first and second IBPGR/ QDPI Banana Germplasm Collecting Mission to Papua New Guinea. Evaluación de impacto del proyecto de banano orgánico en el valle del Chira, Perú En 1997, en el valle del Chira, Perú, se registró la menor superficie cultivada de banano durante los últimos 20 años principalmente debido la baja rentabilidad del cultivo, bajo precio de mercado, que alcanzó el nivel de 0.23 dólares por caja de 18 kg, sistema de comercialización en monopolio, bajos rendimientos derivados de la falta de apoyo en asistencia técnica y falta de créditos, pérdida de producción causada por los efectos del fenómeno El Niño y carencia de otras alternativas para la comercialización. En 1998, mediante un acuerdo entre el Ministerio de Agricultura (MINAG) del Perú y INIBAP, se implementó en el departamento de Piura el proyecto de banano orgánico del valle del Chira con características de tipo participativo, enmarcado a la conservación del medio ambiente y la sostenibilidad. El objetivo del proyecto fue caracterizar la problemática bananera e implementar con el apoyo coordinado entre las dos organizaciones 32 actividades de transferencia de tecnología, apoyo crediticio y logístico, investigación y organización de productores de banano para mejorar el nivel de ingresos de los pequeños productores adoptando el cultivo de banano orgánico como forma de producción con fines de diversificar la oferta, mejorando el sistema de producción y apoyo en la comercialización. El valle del Chira se caracteriza por tener suelos sedimentarios, profundos, de baja pendiente y textura franco arcillosa de clase I aptos para el cultivo de banano. El clima de la región es óptimo para el desarrollo del banano orgánico con temperaturas promedio máxima y mínima de 32°C y 17°C en el verano e invierno respectivamente, heliofanía con 7.2 horas por día que favorecen el desarrollo del banano. La humedad relativa menor a 65% y la baja precipitación (menor a 500 mm por año) no favorecen el desarrollo de la Sigatoka negra. La zona de producción está estratégicamente ubicada a 60 km del puerto de embarque y InfoMusa - Vol. 13 - No.1 A través del proyecto se capacitó a los pequeños productores, técnicos locales tanto de organizaciones e instituciones del estado como del sector privado. La adopción de la producción de banano orgánico redujo el uso de fertilizantes de origen sintético en 18 018 TM, los cuales fueron reemplazados por fuentes nutritivas de origen orgánico como el guano de las islas, estiércol de ganado, humus de lombriz, sulfato de potasio y magnesio mineralizado y otras fuentes (Figura 1). Los beneficiarios del proyecto fueron 1672 pequeños productores capacitados, los cuales representaron el 38% de la totalidad de productores de banano en el valle del Chira. A finales de 1999 se adoptó la tecnología en 1603 hectáreas, lográndose certificar con la agencia OCIA-USA las primeras 210 hectáreas de banano que cumplieron con los más exigentes estándares de la regulación de productos orgánicos. En el 2000 se exportó la producción de 115 hectáreas en forma efectiva. En el 2002, se adoptó la tecnología de producción orgánica en cerca de 3100 hectáreas de las cuales 1600 fueron certificadas por las agencias BSC, Skal y SGS. De esta cifra, la producción de 823 hectáreas fueron para la exportación efectiva a través de las empresas nacionales Gronsa, Exbanor y Biorgánika, y la transnacional Dole. El cambio de tecnología produjo modificaciones importantes en el nivel de ingreso tanto de los pequeños productores de banano orgánico como de los productores convencionales quienes se han favorecido de la derrama económica generada por el proyecto. El precio real que recibió el productor por el banano orgánico de exportación en el 2002 fue 32% superior al precio real que InfoMusa - Vol. 13 - No.1 Figura 1. Plantaciones de banano orgánico certificado en el valle del Chira. Figura 2. Precios reales al productor. 30 US$ por milles de frutas Logros alcanzados recibió en 1999 por la venta en el mercado nacional (Figura 2). El costo de producción del banano orgánico por hectárea fue mayor en 92% al costo de producción convencional (Figura 3). Los componentes que determinaron el elevado costo de la producción orgánica fueron la mano de obra y fertilizantes. En 1998, antes de iniciar el proyecto de banano orgánico, el ingreso neto promedio por la venta de banano fue US$963/ha (Figura 4). En el 2002, el ingreso neto promedio del productor fue US$2770. El pequeño productor incrementó su nivel de ingresos en 187% por adoptar la tecnología de producción orgánica y diversificar su oferta al mercado interno y externo. El ingreso neto real promedio por hectárea disminuyó en 65% en el proceso de transición de convencional a orgánico. Este efecto se debió al incremento del costo de producción y a que los precios del banano en transición en el mercado interno no cambiaron con relación al banano convencional. Esta situación se revertió a partir del 2000, en donde la producción orgánica fue exportada a precios diferenciales y el precio real en el mercado nacional subió considerablemente. El efecto 25 20 15 10 Orgánico Convencional 5 0 1999 2000 2001 2002 Figura 3. Costos reales de producción. 2000 US$ por hectárea cuenta con sistemas viales asfaltados con acceso directo. El 98% de los productores del valle del Chira tiene en promedio 0.72 hectáreas. El objetivo del presente estudio fue evaluar ex post el impacto económico de la implementación del proyecto. El estudio de campo se realizó entre octubre del 2002 y marzo del 2003, utilizándose las metodologías de aplicación de 175 encuestas directas a productores, trabajo en grupos de enfoque, reuniones individuales con comerciantes, obtención directa en campo de información de datos técnicos, productivos y de comercio, y recopilación bibliográfica y estadística de información referente a la actividad bananera en la región. El muestreo realizado fue al azar considerando productores orgánicos y convencionales. 1000 0 Orgánico Convencional 1999 2000 2001 33 2002 US$ por hectárea 3000 2000 1000 Orgánico 0 Convencional 1999 2000 Figura 4. Ingresos netos reales. Este trabajo fue realizado por Salomón Soldevilla Canales que trabaja en el Proyecto Banano Orgánico Alto Beni, Sopocachi, Calle Francisco Bedregal N° 2904 P. B., La Paz, Bolivia In memoriam 2001 2002 de la transición a la producción orgánica puede ser más prolongado y de mayor perjuicio para el productor con relación a las condiciones iniciales con las que cuenta el productor involucrado en la conversión del proceso de producción. En el caso de los productores del valle del Chira, la recuperación del ingreso neto real se produjo a los 15 meses de haber iniciado el proceso de conversión, toda vez que los productores, antes de entrar a este proceso de tres años adoptado por la reglamentación de productos orgánicos, ya tenían un período de dos años utilizando prácticas ecológicas o no contaminantes y, en algunos caso, más de tres años sin uso de productos químicos prohibidos. La reducción de la oferta de banano para el mercado nacional causado por la exportación provocó incrementos del precio recibido por el productor, sin que represente necesariamente un incremento del precio al consumidor. Asimismo, los productores convencionales se beneficiaron del cambio de la dinámica de comercialización del banano y la apertura del mercado exterior. En tal sentido, el productor convencional elevó su ingreso neto real en el año 2002 en 107% en comparación a los ingresos registrados en 1998 debido principalmente al mejor precio en campo. En el 2002, los productores orgánicos certificados no pudieron exportar el 24% de la producción orgánica obtenida para la exportación por dificultades en el sistema de comercialización de las empresas exportadoras (incumplimiento de compras según fechas acordadas). El 61% de la cantidad no exportada cumplió con los requisitos mínimos para la exportación y se destinó al mercado nacional por el cual el productor recibió precios reales inferiores del orden del 74% del precio que hubieran recibido de haber exportado. Perspectivas El cambio de sistema de producción y el apoyo a la comercialización promovido por el convenio MINAG-INIBAP contribuyó a que el banano, de ser el 17o frutal más exportado en 1999, pasara a ocupar, en el 2002, el tercer lugar con un valor de exportación de 6.1 millones de dólares seguido del mango y mandarina. El banano registró el mayor crecimiento tanto en el valor como en volumen de exportación. La producción de banano orgánico en el valle del Chira seguirá creciendo en la medida que la demanda por productos orgánicos certificados en el exterior aumente y las empresas nacionales o extranjeras mejoren el sistema de producción, de operación y seguimiento a fin de garantizar un precio justo al productor y la integridad orgánica de la producción. Del mismo modo, las perspectivas pueden mejorar de haber una participación del Gobierno en sectores estratégicos como son la investigación en campo y de mercados, liberalización de la producción y comercio del guano de las islas, fomento para reducir los costos en el transporte marítimo, mejoramiento de vías de acceso y establecimiento de normas claras para estimular la inversión y la distribución de los beneficios generados. Tributo a Georges F. Wilson El Dr Georges F. Wilson, pionero de la investigación moderna de plátanos en Africa, falleció el 7 de marzo en Kingston, Jamaica, después de una prolongada enfermedad. El Dr Wilson desempeñó un papel importante en la creación del programa de banano y plátano en el IITA y de INIBAP. Lanzó sus primeros ensayos con el plátano en Nigeria a principios de la década de los 34 70, cuando se desempeñaba como Oficial Principal del Programa de Investigación de los Sistemas de Cultivo del IITA. Percibió la importancia vital del cultivo en Africa mucho antes de que este fuera reconocido como tal por el IITA y trabajó incansablemente para que los científicos africanos unieran sus esfuerzos de investigación. InfoMusa - Vol. 13 - No.1 Estuvo detrás de la Primera Conferencia Internacional sobre el Banano y el Plátano celebrada en 1976 en la sede del IITA con el apoyo de la Agencia Belga para la Cooperación en Desarrollo. Participaron más de 50 investigadores y especialistas africanos, de todo el mundo. Se estableció una Asociación Internacional para la Investigación del Plátano y otros Bananos de Cocción (IARPCB), con el difunto Jean Champion como primer Presidente, el difunto Harry Stover como Vicepresidente y Georges como secretario. Gracias a Georges, el concepto de una red de investigación bananera se convirtió en realidad. La Corporación para la Investigación de Plátanos de Africa Occidental (WARCORP) fue creada con el apoyo del IFAD y reunió a científicos desde Congo hasta Guinea. La Subestación High Rainfall del IITA en Onne desempeñó un papel esencial en esta red. Sólo unas pocas personas sabían que Georges puede ser considerado como uno de los padres fundadores de esta estación. Durante la 2a conferencia de IARPCB celebrada en Abidjan, Côte d’Ivoire, la llegada de la Sigatoka negra al Africa generó muchas discusiones, que llevaron a la idea de crear una red global para manejar esta nueva y muy seria amenaza. La idea fue apoyada por el Centro Internacional de Investigación para el Desarrollo y Barry Nestel como su consejero principal, y llevó a la creación de INIBAP. Georges tenía una personalidad única, una fuerza silenciosa que lograba sus metas con una gran sabiduría y modestia. Su oficina siempre estaba abierta para todos a cualquier hora del día. Compartía generosamente sus vastos conocimientos en agricultura con las generaciones más jóvenes. En colaboración con su amigo Edmond De Langhe, continuó el desarrollo de la investigación africana en Musa. Nombró a Rony Swennen como primer científico en plátanos en la estación de Onne y le solicitó recolectar los bananos y plátanos de todos los lugares. Esta actividad condujo a la creación del Centro de Tránsito de INIBAP. Con el nombramiento del difunto Dirk Vuylsteke, se trajo al primer biotecnólogo de banano al IITA. Dirk y Georges crearon los primeros centros de multiplicación de plátano in vitro en el IITA, con la asistencia del Banco Mundial. Georges ayudó a estos jóvenes científicos a convertirse en expertos en banano y plátano, quienes compartieron su amor por Africa. Les enseñó que no existen problemas, sólo soluciones. Lo que es más admirable, permaneció siendo un optimista y nunca se rindió. También disfrutó completamente la vida con su esposa Peju e hijo Suen. Siempre le recordaremos sobre todo como El Jamaiquino con el bigote de Clark Gable, como lo describió el Washington Post destacando sus logros en la investigación de plátanos en 1984. Edmond De Langhe y Rony Swennen Mapeo genético y citogenético en bananos: caracterización de translocaciones Tesis Alberto Vilarinhos Tesis de Doctorado (PhD) presentado ante l’Ecole Nationale Supérieure Agronomique de Montpellier, Montpellier, Francia, marzo de 2004 Frecuentemente en el banano se observan irregularidades durante la meiosis. En Musa acuminata, la especie principal en el origen de los cultivares de banano, se han identificado siete grupos de translocación (Estándar, Norte de Malasia, Montañas de Malasia, Norte A, Norte B, Indonesia, y Africa Oriental). Dentro de cada grupo las accesiones tenían la misma estructura cromosómica. En general, las translocaciones crean un problema para el mapeo genético, el estudio genético de los caracteres agronómicos y el mejoramiento del banano. El objetivo de esta InfoMusa - Vol. 13 - No.1 tesis consistía en establecer una herramienta basada en la hibridación fluorescente in situ de los clones de BAC (BAC-FISH) para caracterizar las translocaciones en el banano y utilizar esta herramienta para caracterizar las translocaciones en ‘Calcutta 4’ (2n=2x=22, grupo de translocación Norte A group) y en ‘Madang’ (2n=2x=22, grupo de translocación Estándar). Para lograr este objetivo, se desarrolló una biblioteca de los BAC de banano, se adaptó la metodología de hibridación in situ de BAC para ser utilizada con los cromosomas de banano y se empezó el desarrollo de un mapa citogenético de banano. La biblioteca de BAC tiene 55 152 clones, con un tamaño de inserción promedio de 100 Kb. Alrededor de 1.5% de estos 35 clones tienen inserciones de cloroplastos y mitocondrias. Esta biblioteca cubre de 9 a 10 veces el genoma del banano. Se construyó un mapa genético del cruzamiento ‘Calcutta 4’ x ‘Madang’ utilizando 120 marcadores (20 marcadores de RFLP, 81 marcadores de AFLP y 19 marcadores SSR) distribuidos a través de 14 grupos de enlace. El grupo de enlace II fue seleccionado para buscar los puntos de quiebre para las translocaciones. Algunas características de este grupo (como el alto número de marcadores distorsionados, el tamaño del grupo y la comparación con otros mapas de banano) sugieren la presencia de las translocaciones. Cuatro clones de BAC distribuidos en el grupo de enlace II y seleccionados de tres loci de RFLP y un locus de SSR fueron localizados en los cromosomas de ‘Calcutta 4’ y ‘Madang’, utilizando BACFISH. Los resultados sugieren que los marcadores del grupo de enlace II están Tesis localizados en tres pares de cromosomas homólogos que se diferencian por la presencia de dos translocaciones enlazadas en ‘Calcutta 4’. De acuerdo a nuestra hipótesis, los marcadores mMaCIR161-rMaCIR 560, rMaCIR 1125 y rMaCIR 36 están localizados en tres cromosomas (cromosomas A, B y C) en ‘Madang’. En ‘Calcutta 4’ estos marcadores están localizados sólo en dos cromosomas (cromosomas A y B). El marcador rMaCIR 1125 localizado en el cromosoma C en ‘Madang’, está localizado en la posición proximal del cromosoma B en ‘Calcutta 4’. Paralelamente, se ha empezado un mapa global citogenético de la accesión ‘Calcutta 4’. Este mapa citogenético es parcial e incluye solo 16 loci (14 clones BAC seleccionados por los marcadores RFLP y SSR y las sondas ribosomáticas 45S y 5S). Se han anclado seis de los 14 grupos de enlace del mapa genético del ‘Calcutta 4’. Cultivo de tejidos de Musa acuminata Colla R. Vidhya Tesis de Doctorado (Ph.D) presentado en 2002 ante la Universidad de Kerala, Thiruvananathapuram, Kerala, India Para cumplir con la creciente demanda por los bananos, se debe incrementar la productividad. Tecnologías modernas, como el uso de las plantas in vitro dieron como resultado una productividad síncrona, rápida y mejorada. Los retoños de espada de los bananos ‘Red banana’ fueron recolectados en los campos bien mantenidos en Vellayani y Kaliyilkkavila en el Distrito de Thiruvananthapuram. Los explantes de las puntas apicales fueron esterilizados dos veces e inoculados en el medio basal MS con carbón activado al 0.2%. Después de 4 semanas, se seleccionaron los cultivos estériles, y las vainas foliares y las puntas apicales fueron aisladas e inoculadas en el medio MS y en el medio MT de varias concentraciones y en varias combinaciones de auxinas y citoquininas. Callos friables de color rojo pálido fueron iniciados a partir de los explantes de puntas apicales en el medio MT con 0.5 mg/L de TDZ y se obtuvieron callos compactos de color rojo en el medio MT con 1 mg/L de NAA, 2 mg/L de BA y 1 mg/L de TDZ. Los explantes de las puntas apicales en el medio MS con 2 mg/L de 2,4-D produjeron callos friables de color blanco. Los explantes de vainas foliares en el medio MT con 1 mg/L de NAA y 2 mg/1 de BA y 1 mg/L de TDZ produjeron callos friables de color rojo. El medio 36 MS con 1mg/L de NAA y 2 mg/L de BA produjo callos de color verde pálido menos compactos. La rizogénesis fue observada en los callos iniciados a partir de los explantes de puntas apicales en el medio MT con 1 mg/L de NAA y 2 mg/L de BA. Los callos iniciados a partir de los explantes de vainas foliares en el medio MT con 1 mg/L de NAA y 1 mg/L de BA o 0.1 mg/L de TDZ también mostraron rizogénesis. Los explantes de vainas foliares en el medio MT con 2 mg/L de 2,4-D y de BA produjeron callos de color negro con embriones somáticos en diferentes etapas de desarrollo como los embriones globulares, de torpedo y bipolares. En el cultivo de suspensión, el medio MT líquido con 1 mg/L de 2,4-D y 2 mg/L de BA produjo el mayor número de embriones somáticos después de 45 días. Los embriones somáticos fueron transferidos al medio basal MT para la siguiente etapa de crecimiento. Los brotes florales masculinos terminales fueron recolectados para otro cultivo de plantas con racimos. De las anteras se iniciaron callos esponjosos blancos después de 60 días en el medio MT con 2 mg/L de 2,4-D y de BA; de 2 mg/L de 2,4-D y 1 mg/L de TDZ se iniciaron callos esponjosos de color amarillo pálido y en 2 mg/L de NAA y 1 mg/L de BA, fueron producidos callos negros con raíces. Los explantes de las puntas apicales fueron inoculados en el medio MS para la regeneración directa y multiplicación. Los brotes primarios emergieron de los explantes dentro de una InfoMusa - Vol. 13 - No.1 semana, independientemente de la concentración de hormonas. Múltiples brotes fueron producidos 60 días después de la inoculación, y 78 y 72 inicios de brotes, con un largo promedio de 6.95cm y 6.13 cm respectivamente, se obtuvieron en el medio MS complementado con 8mg/L de BA solo o con 0.1 mg/L de ANA y 8 mg/L de BA. Las plántulas in vitro de los frascos de cultivo fueron aclimatadas y transferidas del cuarto de cultivo a temperatura ambiente. Las plantas regeneradas mostraron un 100% de supervivencia. Todas las plántulas estaban verdes durante la fase inicial de aclimatación. Pero 10 días después, la mayoría de los pseudotallos se tornaron rojos, con excepción de 36 plántulas provenientes de 8 mg/L de BA solo o con 0.1 mg/L de ANA, las cuales permanecieron verdes. Caracteres morfológicos como la altura y la circunferencia de la planta, la cantidad de hojas, largo y ancho de las hojas, fueron registrados al comienzo y después de 30, 90, 180 y 270 días. Las plantas rojas produjeron un promedio de 80 a 90 frutas y las plantas verdes produjeron frutas de tamaño similar y se tornaron amarillas durante la maduración. Se registraron el largo y la circunferencia de la fruta, el peso de la fruta, el peso del racimo, la cantidad de manos, la cantidad de dedos por mano y el número total de frutas de las plantas regeneradas obtenidas de diferentes concentraciones de hormonas y establecidas en cuatro ecotipos bajo 15 tratamientos, y luego se realizó un análisis estadístico. Las muestras de hojas de las plantas obtenidas del medio complementado con 8 mg/L de BA o Kin y establecidas en el campo y las variantes rojas y verdes obtenidas de las plantas in vitro fueron utilizadas para el SDS-PAGE con el fin de separar las proteínas. Se analizaron isozimas como la esterasa, la fosfotasa ácida y la peroxidasa, y los patrones de las isozimas en las plantas regeneradas. El electroforograma de la esterasa y peroxidasa fueron diferentes para las muestras de las hojas de las plantas derivadas de BA y Kin. El electroforograma de la peroxidasa mostró 5 bandas principales para las muestras foliares de Kin, pero solo 3 bandas se obtuvieron para las plantas derivadas de BA. Todas las proteínas en el experimento en gel mostraron variabilidad en el patrón de las bandas de la proteína total. El banano, una fruta tropical importante, proporciona una buena fuente de carbohidratos, vitaminas y minerales. Por mucho tiempo esta fruta fue considerada como el alimento infantil ideal. El contenido de carbohidratos fue más bajo en las variantes rojas y verdes (22.98 y 23.12 mg/g) que en los cultivares locales ‘Robusta’, ‘Nendran’ y ‘Rasthali’. El contenido de vitamina C fue ligeramente más alto en la variante roja. Los niveles más bajos de azúcares reductores fueron obtenidos de las variantes rojas. No hubo diferencias en el contenido nutritivo de las frutas de las variantes rojas y verdes y en el contenido de fibra en el pseudotallo de las muestras derivadas de BA o Kin. Las variantes roja y verde fueron ricas en NDF, ADF, contenido de hemicelulosas y lignina. Se realizó un análisis RAPD con 20 iniciadores aleatorios de Operon Technologies. De estos, 10 iniciadores dieron productos de amplificación. Se anotó un total de 96 bandas, de las cuales 79 bandas eran polimórficas (83%). Después de tres semanas en vermiculita, las variantes roja y verde mostraron polimorfismo marcado en las bandas principales. El OPB-13 puede ser utilizado como un marcador para identificar variantes rojas y verdes obtenidas durante el cultivo in vitro de Musa acuminata. El OPAB-13 puede ser utilizado para distinguir las variantes rojas y verdes. El OPB-3 mostró un patrón de bandas monomórfico para todas las muestras rojas y verdes. De acuerdo al dendrograma UPGMA, las variantes rojas y verdes estaban concentradas en grupos separados. Insumos biotecnológicos para mejorar el rendimiento del banano Niteen V. Phirke Tesis de Doctorado (PhD) presentada ante la Universidad de North Maharashtra, India, febrero de 2002 El principal objetivo de este trabajo fue desarrollar un régimen de bajo costo para el mejoramiento cualitativo y cuantitativo de un sistema de producción de bananos que no comprometiera la fertilidad del suelo, los InfoMusa - Vol. 13 - No.1 intereses de los agricultores y consumidores de banano y la salud del ecosistema. En los sitios de ensayo de la Universidad de North Maharashtra y de la finca agrotecnológica de Bajirao se estableció un diseño de bloques aleatorios con el fin de evaluar el efecto de varios insumos biotecnológicos, como la aplicación del acondicionador del suelo derivado del pseudotallo, fitoreguladores de crecimiento, 37 Tesis biofertilizantes, cenizas volátiles, riego por goteo y una menor cantidad de fertilizantes químicos, sobre el crecimiento y rendimiento del ‘Shrimanti’ (AAA). Los principales resultados de los cinco años de experimentos en el laboratorio y en el campo fueron los siguientes: 1. Aproximadamente 4 millones de toneladas métricas de biomasa de pseudotallo y de hojas por año fueron utilizados para producir un acondicionador de suelo, utilizando fermentación en estado sólido para el carbón orgánico y reciclaje de nutrientes. 2. Los fitoreguladores de crecimiento, basados en aminoácidos producidos hidrolizando subproductos ricos en proteínas disponibles localmente, aumentaron la tasa supervivencia de las plantas transplantadas y la productividad del banano. 3. Microbios eficientes fueron aislados de la rizosfera de las plantas de banano élites y conservados para la explotación comercial en un consorcio de biofertilizantes. Noticias de Musa Figura 1. Lesiones producidas por Verticillium (Stachylidium ) theobromae (Turc.) 4. Las cenizas volátiles mostraron potencial como un sustituto parcial para los fertilizantes fosfatados y potásicos y los micronutrientes importados, conjuntamente con los hongos y micorrizas que solubilizan fosfatos. 5. El riego por goteo redujo la cantidad de agua utilizada, consumo de electricidad y, como resultado, salinización del suelo. 6. Utilizando estos insumos biotecnológicos, la utilización de los fertilizantes químicos se redujo en un 50%, lo que hizo posible la conversión del terreno muy erosionado, árido y sin utilización, en un terreno agrícola cultivable. 7. Los ensayos en dos condiciones geoclimáticas en la Universidad de North Maharashtra y en la finca agrotecnológica de Bajirao han mostrado ampliamente la misma tendencia en las observaciones y productividad, indicando la confiabilidad de la tecnología de manejo integrado de la nutrición de las plantas. Punta de cigarro en Cuba La conocida punta de cigarro (tabaco) es una enfermedad que se caracteriza por la aparición de una zona negruzca en el perianto que se disemina hacia abajo, tornándose corrugado y cubierto de conidióforos y conidios pulverulentos del patógeno, de donde toma el nombre el nombre. En Cuba, esta enfermedad es más frecuente en los llamados plátanos de cocción o viandas, siendo los clones ‘Macho 3⁄4’ y ‘CEMSA 3⁄4’ los más afectados. En áreas del Instituto de Investigaciones en Viandas Tropicales, ubicado en Santo Domingo en la provincia de Villa Clara, se observaron en racimos del clon FHIA-01 (AAAB) un manchado de los frutos que se manifestaba por la aparición de lesiones negruzcas hundidas con bordes sinuosos o irregulares, deprimidos en el centro, de aspecto o consistencia acuosa (Figura 1). La fusión de estas lesiones producen áreas continuas que abarcan hasta las dos terceras partes del dedo. Las lesiones se circunscriben a la corteza, pues la pulpa debajo de ésta no se afecta. Se aprecia decoloraciones pardo rojizas en el pedúnculo de 38 los dedos que pueden llegar a extenderse hasta la inserción con el tallo principal. No se producen pudriciones ni deterioro en los dedos, ya que estos permanecen firmes hasta la maduración que retarda más allá de lo habitual. La parte cóncava del dedo se ve más afectada que la convexa, debido presumiblemente a la difusión que realiza el rocío o las lluvias de las estructuras del patógeno. Se colocaron varias secciones del tejido afectado en cámaras húmedas para favorecer la emisión de estructuras fungosas y se incubaron a 28°C, luego de cuatro días de incubación se detectaron sobre las lesiones un micelio escaso pardo claro, que al observarse al microscopio compuesto, se pudo identificar dichas estructuras como pertenecientes a Verticillium (Stachylidium) theobromae (Turc.). Este hongo se caracteriza por producir conidióforos solitarios, erectos de 100–400 nm x 4–6 nm, septados, cilíndricos, amarillos claro con ramas verticiladas en cuyos extremos se forman masas de conidios embebidos en un mucílago, hialino, oblongos o cilíndricos de 4-6 nm x 2 nm. Fuente: Lilián Morales Romero, Instituto de Investigaciones en Viandas Tropicales. (INIVIT) and Lidcay Herrera Isla, Facultad de Ciencias Agropecuarias, UCLV, Cuba. InfoMusa - Vol. 13 - No.1 Apuntalando las plantas de plátano caídas Noticias de Musa Plantas de plátano dañadas y desarraigadas fueron apuntaladas en una finca en Ipaja Lagos, Nigeria. Puntales en forma de Y fueron utilizados en varios puntos a lo largo del pseudotallo. En 1997, una planta apuntalada a la altura de 90 cm sobre el nivel de la tierra a 45° produjo retoños, pero las raíces fueron comidas por las termitas después de una sequía y la planta se cayó bajo su peso. Otra planta apuntalada resbaló de su soporte después de ocho semanas y cayó sobre una cerca con un ángulo de 65°. El pseudotallo se marchitó en la semana 12 y produjo 10 dedos en la semana 18. En 1998, una planta de plátano fue apuntalada verticalmente, a 80°. Ella empezó a crecer 18 meses después. El largo del pseudotallo aumentó de 165 cm a 180 cm y casi un mes después la planta produjo 3 dedos. La circunferencia del pseudotallo permaneció sin cambios en 37.5 cm. En 2001, dos plantas se cayeron sobre una cerca con inclinaciones de 40° y 60°. La planta apuntalada a 60° produjo un retoño mientras que la otra se marchitó como todas otras plantas que no fueron apuntaladas. Para más información, Godwin Norense Osarumwense Asemota, Windhoek, Namibia [email protected] Jean Champion y la creación de INIBAP Desarrollo de Canadá (CIID) realizar un estudio sobre el alcance y factibilidad de desarrollar alguna forma de apoyo a la investigación de los bananos y plátanos bajo el paraguas del CGIAR. El CIID contrató a un consultor para discutir las posibles opciones con los países productores y donantes potenciales y su informe fue presentado por el CIID en una reunión asistida por 16 donantes en Washington en noviembre de 1983. La reunión llegó a un consenso de que sería apropiado crear alguna forma de iniciativa internacional para apoyar el mejoramiento de los bananos y plátanos y que esta iniciativa debería ser implementada más bien a través de un enfoque de trabajo en una red, que creando un gran instituto multidisciplinario en cualquier lugar. Se le solicitó al CIID realizar más consultas con los programas nacionales de investigación bananera, especialistas y agencias donantes y presentar una propuesta formal al CGIAR en su siguiente reunión en mayo de 1984. Uno de los principales elementos en este proceso de consulta fue una reunión que se celebró en el Aeropuerto de Gatwick, en diciembre de 1983, cuando un grupo de cuatro expertos en Musa a nivel mundial se reunió con el consultor del CIID para ofrecer sus puntos de vista sobre algunas cuestiones clave que habían surgido en los países durante el proceso de consultas regionales. Los tópicos destacados cubiertos identificaban los vacíos más importantes en el conocimiento técnico con respecto a la producción de los bananos como alimentos, sugiriendo prioridades globales y regionales para la investigación, El Dr Jean Champion, de cuyo fallecimiento hemos informado en la edición de diciembre de 2003 de INFOMUSA, desempeñó un papel poco conocido pero significativo en la creación de INIBAP. Formó parte de un pequeño grupo sin cuya experiencia y entusiasmo la red nunca podría haber llegado a existir. A principios de la década de los 80, los miembros del Grupo Consultivo para la Investigación Agrícola Internacional (CGIAR) estaban discutiendo la creación de varios nuevos Centros de Investigación Agrícola Internacional (IARC). Varios países productores de bananos y plátanos sugirieron que Musa representaba el cultivo alimentario más importante y el cual no recibía atención del CGIAR. En este tiempo, la mayoría de los países donantes pensaban que los bananos constituían principalmente un producto de exportación comercializado por las compañías multinacionales, y la importancia de los bananos y plátanos como un producto alimentario en los países en vías de desarrollo no estaba reconocida completamente. Esta situación empezó a cambiar cuando la enfermedad de la Sigatoka negra, que tuvo un impacto devastador sobre los pequeños productores, se propagó rápidamente por todo el mundo. A finales de 1982, un grupo de los principales donantes en una reunión en Washington solicitó al Centro Internacional para la Investigación y InfoMusa - Vol. 13 - No.1 Una planta caída apoyada por luna cerca (Goodwin Asemota). 39 Forum desarrollando una estrategia internacional para el mejoramiento y discutiendo las necesidades de la tecnología de información del nuevo instituto, necesidades de capacitación y la estructura óptima de una red internacional. El Dr Champion desempeñó un papel clave en esta consulta, ya que sus descubrimientos establecieron el marco tanto para la aprobación de la creación de INIBAP por el CGIAR en mayo de 1984, como para la definición de las políticas y programas aprobados por la primera Junta directiva de INIBAP. Antes de la creación formal de INIBAP, fue necesario identificar un país anfitrión para su sede. Esta fue una decisión difícil ya que la producción de Musa estaba dividida casi igualmente entre las regiones de Africa, Asia y América Latina y el Caribe, pero ninguna de las regiones tenía un fuerte programa de Musa, aparte del grupo de América Central perteneciente a una corporación multinacional. El centro nacional de excelencia más fuerte fue el CIRAD (Centre de coopération internationale en recherche agronomique pour le développement), con su personal en Montpellier y en otros países. Algunos donantes estaban preocupados sobre la ubicación del instituto líder para un cultivo tropical en Europa y tenían dudas sobre el modelo propuesto de trabajo en red. Esta situación llevó a más diálogos y consultas. Tomando en cuenta al CIRAD, donde Jean Champion fue uno de los principales y más respetados científicos, y varios países productores que apoyaban la candidatura de Montpellier, el Gobierno Francés ofreció esta ciudad para la sede de INIBAP. El resto, como se dice, es historia. Barry Nestel, Consultor Si, hubo bananos en Camerún hace más de 2000 años La llegada de los primeros bananos cultivados al Africa ha sido una cuestión de conjeturas por más de un siglo. Mientras se acepta comúnmente que ellos se originaron en “algún lugar del Este”, no se ha determinado con seguridad cuando ellos fueron introducidos y quien fue responsable. Había tres teorías para explicar la introducción de los bananos al Africa. Los bananos fueron introducidos por los portugueses al final del siglo 16, por los comerciantes árabes o persas alrededor del siglo 8o o antes, o por las personas que hablaban el idioma austronesio, quienes se habían establecido en Madagascar a principios del primer milenio, haciendo posible una introducción posterior al continente. La tercera teoría, desarrollada por el difunto Norman Simmonds, una autoridad en bananos, 40 ha estado ganando terreno firmemente. Común a todas las teorías, es la creencia de que los bananos no llegaron al continente africano antes de la era cristiana, hace 2000 años. Recientemente, hemos recuperado fitolitos de los pozos de deshechos excavados en Camerún central e identificado como provenientes de un banano cultivado, después de un estudio comparativo del género Musa y Ensete (Mbida et al. 2001). Estos fitolitos databan de antes de 2500 años antes de Cristo. Si se confirma, este descubrimiento arrojaría una luz diferente sobre la evolución temprana de la agricultura en los trópicos húmedos de Africa. Por ejemplo, la agricultura en los bosques húmedos contaría no solo con el ñame, el cual generalmente no es muy productivo en ausencia de una estación seca, sino que el mismo podría haberse desarrollado alrededor de los plátanos, los cuales prefieren este ambiente. La determinación del primer cultivo de bananos en Africa es muy relevante para la genética de los bananos comestibles. Los plátanos AAB y los bananos de cocción AAA de Africa Oriental comprenden grupos de cultivares estrictamente definidos, que están clasificados morfológicamente en el mismo nivel que el AAA Gros Michel, el AAB Silk y el ABB Pisang awak, por ejemplo. Incluso ellos, como el grupo AAB Maia maoli del Pacífico, despliegan una diversidad que excede la de cualquier grupo comparable de triploides, con más de 50 AAA de Africa Oriental y más de 100 cultivares de plátano. ¿Fue tal diversidad generada a través de las mutaciones somáticas durante largos períodos de tiempo o son estos triploides más susceptibles al tipo de estrés que generan estas mutaciones? El hecho de que las tasas de mutación en el laboratorio son mucho más altas que las mutaciones en el campo (Vuylsteke et al. 1991), destaca la presencia y cultivo prolongados de plátanos en Africa. Es inevitable que los datos que apoyan la idea revolucionaria de un antiguo cultivo de bananos en Africa serán examinados críticamente. En una nota publicada en la edición de 2004 de Azania, la revista del Instituto Británico en Africa Oriental, Jan Vansina, una bien conocida autoridad en historia africana, expresa serias reservas sobre estos descubrimientos (Vansina 2004). El escribe que “sólo se puede aceptar que la evidencia más temprana en Africa del cultivo de bananos comestibles sin semillas se remonta a finales del siglo sexto de la era cristiana y posiblemente hasta más tarde, al siglo noveno de la era cristiana.” Vansina sostiene que los bananos podrían no haber sido cultivados en Africa Occidental 2500 años antes de Cristo, basándose en la hipótesis de que los plátanos AAB se originaron en el subcontinente de la India. Sin embargo, la InfoMusa - Vol. 13 - No.1 investigación quimiotaxonómica y el análisis de ADN citoplásmico demuestran que los plátanos AAB (así como los bananos de cocción AAA de Africa Oriental) se originaron en Nueva Guinea y en las islas circundantes (Horry 1989, Lebot et al. 1993, Carreel 1994, Carreel et al. 2002). Además, la presencia en Asia de sólo unas pocas variedades de plátano y la ausencia de los bananos de cocción AAA de Africa Oriental, hace claro que el subcontinente de la India no puede estar en el origen de la diversidad única encontrada en los trópicos húmedos de Africa. Vansina parece no conocer otro camino para la introducción (De Langhe y de Maret 1999), lo que hace que el área más probable para la introducción de los bananos sea el Africa en la zona ecuatorial, como la actual Tanzania. Desde allí los bananos podrían haber sido dispersados a través del continente desde el oriente hasta el occidente, llegando eventualmente al Camerún actual. Las conjeturas sobre el cultivo temprano de bananos en la parte más al norte del continente se vuelven irrelevantes. Vansina considera que los fitolitos encontrados en Camerún pertenecen más bien al género Ensete, el llamado banano africano falso, que a Musa. En su artículo, manifiesta que “no se hizo ninguna comparación directa en el laboratorio con los fitolitos utilizados en los estudios anteriores especialmente en el Sudeste de Asia” (Vansina 2004). Cuando presentamos nuestro trabajo (Mbida et al. 2001), sabíamos sólo de dos estudios de relevancia general para las Musaceae (Tomlinson 1959, Tomlinson 1969) y un estudio específico sobre Nueva Guinea (Wilson 1985). Referimos a estos trabajos, así como a las colecciones de referencia publicadas para los continentes americano (Piperno 1988), asiático (Kealhoffer y Piperno 1998) y africano (Runge 1996, Runge 1997). El trabajo de Wilson no apoya la afirmación de Vansina que “es prácticamente imposible distinguir entre los fitolitos de Musa (bananos) y otras Musáceas (en este caso Musa ingens)” (Vansina 2004). Aún si Wilson encontró difícil de discriminar entre M. ingens y otras secciones de Musa, él aparentemente no introdujo el género Ensete en el estudio1. Más aún, la diferenciación de los fitolitos de Ensete de cualquier fitolito de Musa fue el punto de nuestro estudio (Mbida et al. 2001). Incidentalmente, las indicaciones de Wilson sobre el cultivo temprano de los bananos fueron confirmadas recientemente por Denham et al. (2003), y la domesticación de los bananos puede haber sido iniciada en Kuk (Papua Nueva Guinea) 10 000 años antes de Cristo. La crítica de Vansina que “el material comparativo utilizado es muy limitado”, parece reflejar una interpretación equivocada de los requerimientos metodológicos para InfoMusa - Vol. 13 - No.1 este tipo de estudio específico. Ya que los fitolitos encontrados en Nkang podrían haber pertenecido a las especies africanas nativas de Ensete, se emprendió un extenso estudio comparativo para investigar si la morfología de los fitolitos de Ensete podría ser distinguida de la morfología de los fitolitos del género Musa. Se examinaron varias muestras de Ensete gilletii y Ensete ventricosum, las únicas especies africanas2. Ya que el E. gilletii es típico del paisaje camerunense, un espécimen de una planta que crece allí fue introducida en la colección de referencia, en adición al espécimen de la Colección Internacional de Germoplasma de Musa. No se observó variación notable en la forma del fitolito entre las muestras de Ensete examinadas. Por otro lado, los fitolitos de Musa, si estuvieron presentes en Africa en tiempos antiguos, señalarían necesariamente una introducción de las plantas de banano desde afuera del continente. Los cultivares introducidos podrían haber pertenecido a cualquiera de los genomas de los cultivares de banano, es decir, AA, AAA, AAB y ABB. Los cultivares representativos de estos grupos genómicos fueron examinados con respecto a sus fitolitos. Rechazamos la discrepancia de que el material de referencia fue limitado. Un examen cuidadoso de la morfología de los fitolitos nos llevó a concluir (1) que la variación en la forma de los fitolitos difícilmente se nota dentro de cada género y (2) que la forma ampliamente difiere entre los dos géneros (Mbida et al. 2001). Todas menos una de las características observadas fueron mutuamente excluyentes. Consecuentemente, las dos poblaciones de fitolitos para el Ensete y Musa, respectivamente, son tan distintas que ni siquiera se necesitó un análisis estadístico. Además, la metodología aplicada de observación de los fitolitos no fue criticada por los especialistas o en botánica arqueológica en general, aunque el tópico fue presentado en numerosas conferencias y reuniones desde la publicación de nuestro artículo (Vrydaghs et al. 2003). Resumiendo, mantenemos nuestra conclusión anterior de que los fitolitos del sitio Nkang, que datan de 2500 años antes de Cristo, pertenecen al género Musa y señalan el cultivo de los bananos en esta parte de Africa. Sin embargo, aceptamos que los descubrimientos de los fitolitos en Nkang necesitan ser comprobados por más 1 El Ensete glaucum aparentemente no crece a la altitud de Kuk y los altiplanos circundantes. 2 La distribución de una tercera especie, Ensete homblei, está confinada a una región restringida en Katanga, RD de Congo y Zambia (Simmonds 1960). 41 especimenes, preferiblemente de otros sitios. Esperamos que nuestro debate aliente a más arqueólogos a buscar más fitolitos en los trópicos húmedos de Africa. C.M. Mbida, H. Doutrelepont, L. Vrydaghs y H. Beeckman trabajan en el Royal Museum of Central Africa en Tervuren, Bélgica, R.L. Swennen, R.J. Swennen y E. De Langhe en la Katholieke Universiteit Leuven en Lovaina, Bélgica, y P. de Maret en la University of Brussels en Bruselas, Bélgica. (Autor para correspondencia: rony.swennen@kuleuven. ac.be) Referencias Carreel F. 1994. Etude de la diversité génétique des bananiers genre Musa à l’aide des marqueurs RFLP. Thèse, Institut National Agronomique, ParisGrignon. Carreel F., D. Gonzalez de León, P. Lagoda, C. Lanaud, C. Jenny, J.P. Horry & H. Tezenas du Montcel. 2002. Ascertaining maternal and paternal lineage within Musa by chloroplast and mitochondrial DNA RFLPanalyses. Genome 45:679-698. De Langhe E. & P. de Maret. 1999. Tracking the banana: its significance in early agriculture. In The Prehistory of Food. Appetites for change (Chris Gosden & Jon Hather, eds). Routledge, London and New York. De Langhe E., R. Swennen & D. Vuylsteke. 1994-5. Plantain in the early Bantu world. Azania 29-30: 147-160. Denham T.P., S.C. Haberle, C. Lentfer, R. Fullagar, J. Field, M. Therin, N. Porch & B. Winsborough. 2003. Origins of agriculture at Kuk swamp in the highlands of New Guinea. Science 3010:189-193. Horry J.P. 1989. Chimiotaxonomie et organisation génétique dans le genre Musa. Thèse de Doctorat, Université de Paris XI, France. Kealhoffer L. & D.R. Piperno. 1998. Opal phytoliths in Southeast Asian flora. Smithsonian Contribution to Botany 88. 39pp. Lebot V., K.M. Aradhya, R. Manshardt & B. Meilleur. 1993. Genetic relationships among cultivated Tabla 1. Clasificación de una nueva especie, Musella splendida. Origen Tamaño de la planta Rizoma Relación largo ancho de la hoja Inflorescencia Yema masculina Color de la bráctea Arreglo de la flores Flores básales Forma de la fruta Diseminación Referencia Musella lasiocarpa Provincia de Yunnan, sur de China Pequeña, menos de 60 cm de altura Rizomatoso <3 Musella splendida Provincia de Ha Giang, norte de Vietnam Mediana, 1.0 a 1.2 m de altura No rizomatoso >3 Cónica, deltoidea Marcadamente imbricado, brácteas unidas firmemente a la yema Amarillo, amarillo-naranja Biseriado Femeninas Ovoide Semillas, retoños Wu, D.L. y W.J. Kress. 2000. Musaceae. In C.Y. Wu and P.H. Raven (eds), Fl. China 24:314-318 Ovalada Apice de la yema abierto, brácteas individuales separadas precozmente Amarillo brillante Uniseriado Hermafroditas Avoada Retoños R. Valmayor y L.D. Danh. 2002. Classification and Characterization of Musella splendida sp. nov. Phillip. Agri. Scientist 85(2):204-209 42 bananas and plantains from Asia and the Pacific. Euphytica 67:163-175. Mbida C.M., H. Doutrelepont, L. Vrydaghs, R.L. Swennen, R.J. Swennen, H. Beeckman, E. De Langhe & P. de Maret. 2001. First archaeological evidence of banana cultivation in central Africa during the third millennium before present. Vegetation History and Archaeobotany 10:1-6. Piperno D.R. 1988. Phytolith analysis. An archaeological and geological perspective. San Diego Academic Press. 280pp. Runge F. 1996. Opal phytolithe in pflanzen aus dem humiden und semi-ariden osten Afrikas und ihre bedeutung für die klima- und vegetationsgeschichte. Botanish Jahrb. Systematik 118 (3):33-363. Runge F. & J. Runge. 1997. Opal phytolith in East African plants and soils. Pp. 71-81 in The state-ofthe-art of phytoliths in soils and plants. (A. Pinilla, J. Juan-Tresserras and M.J. Machado, eds). Centro de Ciencas Medioambientales. CSIC Monografias 4. Simmonds N.W. 1960. Notes on banana taxonomy. Kew Bull. 14(2):198-212. Tomlinson P.C. 1959. An anatomical approach to the classification of the Musaceae. J. Linn. Soc. Bot. 55:779-809. Tomlinson P.C. 1969. Anatomy of the Monocotyledons. III. Commelinales-Zingiberales. Oxford, Clarendon Press. Vansina J. 2004. Bananas in Cameroon c. 500 BCE? Not proven. Azania 38:174-176. Vrydaghs L., R. Swennen, C. Mbida, E. De Langhe, H. Doutrelepont & P. de Maret. 2003. The banana phytolith as a direct marker of early agriculture: a review of the evidence. Pp. 177-185 in Phytolith and starch research in the Australian-PacificAsian regions: the state of the art (D. Hart and L. Wallis, eds). Papers from a conference held at the Australian National University, August 2001, Canberra, Australia. Pandanus Books. Research School of Pacific and Asian Studies. The Australian National University. Vuylsteke D., R. Swennen & E. De Langhe. 1991. Somaclonal variation in plantains (Musa spp., AAB group) derived from shoot-tip culture. Fruits 46(4): 429-439. Debate taxonómico En nuestra edición de diciembre de 2002, hemos reproducido un artículo del Philippine Agricultural Scientist, en el cual Ramon Valmayor y Le Dinh Danh anunciaban la clasificación de una nueva especie, Musella splendida R. Valmayor & L.D. Danh sp. nov. Antes de publicar la carta de un lector argumentando la clasificación, presentamos un resumen de las características que los autores utilizaron para diferenciar su nueva especie de Musella lasiocarpa (Franchet) (Tabla 1). InfoMusa - Vol. 13 - No.1 Musella splendida: una respuesta a Valmayor y Danh La prospección de una nueva Musella (Valmayor y Danh 2002) es excitante para los entusiastas de las Musáceas alrededor del mundo, pero mi emoción inicial ha sido apagada. Mi preocupación se centra en el hecho de que los autores basan su separación de Musella splendida en la descripción de Musella lasiocarpa, que ellos asumen cubre toda la especie en todas las situaciones. Ellos se refieren a especimenes de M. lasiocarpa no vivientes o de herbarios e ignoran la posible influencia de los factores edáficos, climáticos y bióticos sobre el crecimiento de M. lasiocarpa. Ellos también ignoran la posibilidad de la variación intraespecífica en M. lasiocarpa y que su material podría caer dentro de este rango. Aunque los autores confían mucho en la altura de la planta para diferenciar una nueva especie, ellos no presentan la base para sus mediciones y ninguna de las fotografías contiene una escala. Los descriptores para el banano de INIBAP especifican que la altura debe medirse desde la base del pseudotallo hasta la punta emergente del pedúnculo. La precisión en materia de la estatura es especialmente importante con Musella, en la cual las hojas están más bien dirigidas hacia arriba y contribuyen así a la altura total de la planta en la fase vegetativa. Sólo alrededor del 30% de la altura pertenece al pseudotallo, el resto son hojas. Tratando con un carácter tan flexible como la altura de la planta, la influencia de las condiciones de crecimiento es crucial. El conocimiento de Valmayor y Danh’s de M. lasiocarpa parece derivar exclusivamente de C.Y. Wu, quien describe una planta “menor de 60 cm de altura”. Valmayor y Danh mencionan, pero no se refieren específicamente a la descripción del tipo de M. lasiocarpa de Franchet (1889) quien también asevera que M. lasiocarpa raramente excede 60 cm de altura. El trabajo de Franchet incluye una ilustración de M. lasiocarpa que posiblemente puede ser una reproducción fiel de la planta descubierta por Abbé Delavay en 1885. Pero el dibujo de Franchet carece de una escala y no se parece a ninguna de las plantas de M. lasiocarpa que yo he visto. De acuerdo al dibujo de Franchet, parece que la altura de la planta, que casi no tiene un pseudotallo, es de 60 cm desde la tierra hasta la punta de la hoja. Baker (1893) estuvo de acuerdo con este hecho y si las mediciones de la altura de Wu derivan, como yo sospecho, de Franchet, entonces en la literatura realmente se indica que M. lasiocarpa es un banano muy pequeño. ¿Pero es esto generalmente verídico para la planta in vivo? Una planta de M. lasiocarpa que crece “sur les rochers de Loko-chan” a 1200 m (Franchet 1889) se verá diferente de una planta que crece en “suelo forestal fértil con humedad abundante” a 118 m en el norte de Vietnam. Al momento de escribir este artículo, la altura desde la base del pseudotallo hasta la punta emergente del pedúnculo de una M. lasiocarpa en fase masculina en mi invernadero (en el sudoeste de RU) fue de 45 cm y la inflorescencia fue de 25 cm por encima. Con hoja totalmente abierta, justamente antes de la floración, mi planta tenía una altura total de 1.3 m, es decir, desde la base del pseudotallo hasta la punta de la hoja más alta en su ángulo natural. M. lasiocarpa puede llegar a ser más alta que esto. Yo envié dos plantas de M. lasiocarpa al Sr Wim Kea de Amstelveen, Holanda. Ellas crecen hasta 2.5 m o más (Figura 1). Yo planteo que M. lasiocarpa es una planta mucho más grande de lo que Valmayor y Danh suponen. Figura 1. Planta de M. lasiocarpa en el jardín del autor. La escala tiene 1 m de largo. Figura 2. a) Inflorescencia de M. lasiocarpa en la fase femenina en el invernadero del autor en junio de 2002. b) La misma inflorescencia en la fase masculina en abril de 2003. c) La misma inflorescencia en la fase masculina en julio de 2003. InfoMusa - Vol. 13 - No.1 43 En la fase femenina, las puntas puntiagudas de las brácteas individuales de M. lasiocarpa pueden realmente estar “separadas precozmente, antes de plegarse hacia la base” (Figura 2a), una característica mediante la cual Valmayor y Danh (2002) diferencian a M. splendida. Pero la característica de las brácteas cambia a medida que la inflorescencia madura, un proceso que en M. lasiocarpa toma meses. En la fase masculina (Figura 2b), las brácteas de M. lasiocarpa son mucho más pequeñas, delgadas y “marcadamente imbricadas”. Este cambio en la característica de las brácteas puede observarse en las propias fotografías de Valmayor y Danh (Figuras 2, 3, 5 y 6 en Valmayor y Danh 2002). Creo que la característica de las brácteas en la fase femenina en M. lasiocarpa puede ser variable y depender de si la planta tiene todas sus hojas o no al momento de empezar la floración. “Los especimenes interesantes de Musella” de Valmayor y Danh (Figura 9, Valmayor y Danh 2002) son, yo planteo, fotografías al azar de M. lasiocarpa. Es prematuro sugerir que la figura 9 representa una evidencia de una posiblemente nueva especie de Musella. Se puede encontrar rápidamente muchas fotografías de M. lasiocarpa en la Internet que muestran aún una mayor variación entre las plantas de la especie. Esta variación principalmente es ambiental o relacionada con la edad de la inflorescencia al momento de fotografiarla. Tampoco estoy seguro que la fruta mostrada en la figura 8 (Valmayor y Danh 2002) pueda ser descrita como partenocárpica. Uno inmediatamente comienza a preguntar mediante que mecanismos esta “especie” se disemina en los “vastos bosques” en el norte de Vietnam. Grandes espacios aéreos visibles en el corte transversal parecen indicar que la fruta no está desarrollada. M. lasiocarpa produce el mismo tipo de fruta como si no fuera polinizada. Valmayor y Danh (2002) manifiestan que M. splendida tiene flores basales hermafroditas y comparan este hecho con M. lasiocarpa que se dice tener flores basales femeninas. Valmayor y Danh no describen propiamente las flores hermafroditas de M. splendida, ni tampoco Wu describe propiamente las flores femeninas de M. lasiocarpa. “Las flores femeninas nacen en la base de la inflorescencia” escribe Wu citado por Valmayor y Danh (2002). Este difícilmente es un diagnóstico. De hecho, la escasez de la publicación de Wu sobre Musella lasiocarpa es una de las razones del porqué la Sociedad Hortícola Real (Royal Horticultural Society) (2003) persiste en referirse a la planta como Musa lasiocarpa, siguiendo a Simmonds (1960). Incidentalmente, Simmonds, quien obviamente no conocía la planta muy bien, basó su 44 inclusión de la planta en Musa en los caracteres del periantio y no en que ella es rizomatosa y policárpica. En mi M. lasiocarpa las flores femeninas tienen filamentos rudimentarios y las flores masculinas, estilos rudimentarios. Es necesario describir propiamente las flores hermafroditas de M. splendida ya que las flores de las Musáceas pueden ser no solo estructuralmente hermafroditas sino también funcionalmente hermafroditas, femeninas, masculinas y hasta estériles. Se puede argumentar que el término debería ser restringido a las flores que son funcionalmente hermafroditas, es decir, que pueden ser autosuficientes para producir semillas viables, como en Musa velutina. Si Valmayor y Danh están utilizando el término en este preciso sentido entonces cual es la explicación para que las flores hermafroditas de M. splendida no produzcan las semillas? Valmayor y Danh no hacen ningún comentario sobre esto o sobre la presencia o viabilidad de cualquier polen producido por estas flores hermafroditas. En conclusión, creo que es prematuro afirmar el descubrimiento de una nueva especie de Musella de Vietnam cuando las poblaciones de Musella en China, Laos, Myanmar y Vietnam están caracterizadas de manera tan inadecuada. Es posible que existan otras especies de Musella esperando ser descubiertas, pero sobre la evidencia que ellos presentan, Valmayor y Danh no me convencen que M. splendida es una de ellas. Tengo una M. lasiocarpa de Kunming, Yunnan, cortesía del Prof. Hu Zhihao, y he depositado cultivos de ápices in vitro de la misma en el Centro de Tránsito de INIBAP en Lovaina. Con mucho gusto intercambiaré material in vivo con cualquier persona interesada en comparar el material uno al lado del otro en el mismo ambiente. Estoy seguro que es interesante desde el punto de vista botánico y posiblemente hortícola la variación útil en M. lasiocarpa. David Constantine 2 High Street Ashcott, Somerset, TA7 9PL, RU e-mail: [email protected] Referencias Baker J.G. 1893. A synopsis of the genera and species of Museae. Ann. Bot. 7:189-229. Franchet A.R. 1889. Un nouveau type de Musa. Musa lasiocarpa. Journ. de Bot. (Morot). 3(20):329-331. Royal Horticultural Society. 2003. RHS Plant Finder. Dorling Kindersley. London. Simmonds N.W. 1960. Notes on banana taxonomy. Kew Bull. 14(2):198-212. Valmayor R.V. & L.D. Danh. 2002. Clasificación y caracterización de Musella splendida sp. nov. INFOMUSA 11(2):24-27. InfoMusa - Vol. 13 - No.1 Instrucciones para los autores INFOMUSA es una revista internacional que se publica dos veces al año en los idiomas inglés, francés y español. Nuestro objetivo es dar a conocer los resultados de investigaciones e informes de interés a toda la comunidad bananera. Ya que INFOMUSA publica artículos sobre cualquier tema relacionado con Musa, los autores deben emplear frases sencillas y cortas para evitar la jerga innecesaria y hacer su trabajo accesible para los lectores, especialistas en otras disciplinas. Los textos mecanografiados deberán ser preparados en inglés, francés o español y no exceder 2500 palabras, incluyendo la bibliografía. El texto debe ser mecanografiado a doble espacio. Todas las páginas (incluyendo las tablas, figuras, leyendas y referencias) deberán estar enumeradas consecutivamente. Incluya el nombre completo de todos los autores y sus direcciones completas al momento de realizar el estudio. También indique la persona quien recibirá la correspondencia respecto al trabajo. El manuscrito debe ser enviado como un adjunto al correo electrónico o en un disquete de 3.5 pulgadas para las computadoras compatibles con PC. Por favor, indique el nombre y la versión del procesador de palabras utilizado y la dirección electrónica del autor. En cualquier caso, necesitaremos recibir por correo dos copias impresas del manuscrito. Título: El título debe ser lo más corto posible y no contener números, siglas, abreviaturas o signos de puntuación. Resúmenes: Es necesario adjuntar un resumen que no exceda de 200-250 palabras. El resumen debe recapitular de manera concisa el contenido básico y enviado en el mismo idioma del manuscrito. Si es posible, se debe enviar las traducciones (incluyendo el título) en los otros dos idiomas. Palabras clave: Facilite un máximo de seis palabras clave, en orden alfabético, debajo del resumen en el idioma del autor. Introducción: La introducción debe proporcionar una razón para la investigación y cualquier información de fondo. Ya que esta introducción no pretende representar una amplia reseña del tópico, la cantidad de referencias bibliográficas debe mantenerse al mínimo. Se debe evitar las introducciones sobre la importancia de los bananos como un producto básico o producto comercial, al menos que sean absolutamente necesarias para el entendimiento del artículo. Materiales y métodos: Los autores deben proporcionar suficientes detalles de su diseño experimental para permitir al lector evaluar la validez de la investigación. Para los materiales y métodos utilizados comúnmente, será suficiente una simple referencia. Resultados: La unidad debe ser separada de la cantidad por un espacio sencillo y seguir la nomenclatura SI, o la nomenclatura común a un campo particular. Las unidades o abreviaturas inusuales deben ser definidas. Presente los datos en el texto o como una figura o una tabla, pero nunca los duplique. Evite el uso extenso de los gráficos para presentar los datos que podrían ser presentados más concisamente en el texto o en una tabla. Limite las fotografías a aquellas que son absolutamente necesarias para mostrar los descubrimientos del experimento. Discusión: La discusión no debe contener repetición extensa de la sección de los resultados, ni reiterar la introducción. Esta parte debe combinarse con la sección de los resultados. Bibliografía: Todas las referencias bibliográficas mencionadas en el texto deberán ser presentadas por el nombre del autor o autores y el año de publicación (por ejemplo: Sarah et al. 1992, Rowe 1995). Se debe evitar referencias a los documentos que circulan ampliamente, como informes anuales, y las citas de las comunicaciones personales y de los datos no publicados. Al final del texto, se debe presentar una lista de la bibliografía en orden alfabético. Por favor, siga el estilo de los siguientes ejemplos: Artículos de ediciones periódicas: Sarah J.L., C. Blavignac & M. Boisseau. 1992. Une méthode de laboratoire pour le criblage variétal des bananiers vis-à-vis de la résistance aux nématodes. Fruits 47(5): 559-564. Libros: Stover R.H. & N.W. Simmonds. 1987. Bananas (3rd edition). Longman, London, United Kingdom. Artículos (o capítulos) en libros: Bakry F. & J.P. Horry. 1994. Musa breeding at CIRAD-FLHOR. Pp. 168-175 in The Improvement and Testing of Musa: a Global Partnership (D.R. Jones, ed.). INIBAP, Montpellier, Francia. Ilustraciones y tablas: Las ilustraciones y tablas deberán estar enumeradas consecuentemente y las referencias en el texto se harán de acuerdo a esta numeración. Cada ilustración o tabla debe incluir un título sencillo y claro. Las figuras y tablas deben insertarse después de la sección de bibliografía o en archivos separados. Gráficos: suministrar los datos correspondientes junto con los gráficos. Dibujos: si es posible, suministrar dibujos originales. Fotografías: Preferimos fotografías de buena calidad (papel brillante con un buen contraste para las películas en blanco y negro, pruebas de buena calidad y películas o diapositivas originales para las fotografías a colores), pero por favor, recuerde que no las devolveremos. Publicaremos las imágenes digitalizadas o tomadas con una cámara digital si su resolución es suficientemente alta (1 millón de píxeles o un mínimo de 300 dpi de una fotografía de tamaño real). Son aceptables los archivos con extensiones JPEG, TIFF y EPS. Evite enviar fotografías insertadas en un documento de Word o Power Point, a menos que estas están acompañadas de una mejor alternativa de calidad. Siglas: Las siglas deberán ser transcritas completamente la primera vez que éstas aparezcan en el texto, seguido por las siglas entre paréntesis. Nombres de cultivares: El nombre del cultivar debe ser colocado entre comillas sencillas. Si el nombre es compuesto, solo la primera palabra empieza con la letra mayúscula, a menos que se refiera a un lugar o nombre de persona. Utilice los nombres comúnmente acordados, como ‘Grande naine’ y evite las variaciones locales o traducciones, como ‘Gran Enano’. Nota: Si el material de plantación utilizado para los experimentos descritos proviene o está registrado en el banco de germoplasma de INIBAP, debe indicarse su número de accesión (código ITC) dentro del texto o en forma tabular. Gracias por seguir nuestras recomendaciones. Esto facilitará y acelerará el trabajo de edición. Publicaciones de INIBAP Nuevas publicaciones S. Mohan Jain y R. Swennen (eds). 2004. Banana improvement, cellular, molecular biology, and induced mutations. Este libro de 392 páginas, co-publicado por FAO/IAEA e INIBAP, presenta los resultados del proyecto de investigación coordinada de FAO/IAEA sobre la biología celular y biotecnología incluyendo técnicas de mutaciones para la creación de nuevos genotipos útiles de banano. El libro también contiene varios artículos de fondo que proporcionan una información actualizada sobre las herramientas biotecnológicas que pueden ser utilizadas para producir, de una manera más rápida y eficiente, nuevas variedades de Musa con caracteres deseables. Publicaciones recientes Strosse H., R. Domergue, B. Panis, J.V. Escalant y F. Côte. 2003. Suspensiones de células embriogénicas de banano y plátano (A. Vézina y C. Picq, eds). Guías técnicas INIBAP 8. INIBAP, Montpellier, Francia. Carlier J., D. De Waele y J.V. Escalant. Evaluación de la resistencia de los bananos al marchitamiento por Fusarium, enfermedades de las manchas foliares causadas por Mycosphaerella y nematodos a escala mundial. Evaluación de comportamiento (A. Vézina y C. Picq, eds). Guías técnicas INIBAP 7. INIBAP, Montpellier, Francia. Pronto disponible INIBAP 2004. Networking bananas and plantains: Annual Report 2003. www.inibap.org Para obtener una lista completa de las publicaciones de INIBAP, visite nuestro sitio web o contacte a Leila Er-rachiq en la sede de INIBAP en Montpellier. Correo electrónico : [email protected] Direcciones de INIBAP • Sede Parc Scientifique Agropolis II 34 397 Montpellier Cedex 5 - Francia E-mail: [email protected] Fax: (33) 467 61 03 34 Director: Dr Richard Markham E-mail: [email protected] Coordinador, Mejoramiento genético de Musa: Dr Jean-Vincent Escalant E-mail: [email protected] Coordinador, Genómica de Musa y conservación de germoplasma: Dr Nicolas Roux E-mail: [email protected] Coordinador Sistemas agroecológicos de Musa y canales de valor agregado: Dr Charles Staver E-mail: [email protected] Coordinadora, Información y Comunicación: Claudine Picq E-mail: [email protected] Coordinadora MGIS: Elizabeth Arnaud E-mail: [email protected] Contable: Emmanuel Gonnord E-mail: [email protected] • Red Regional para América Latina y el Caribe Coordinator Regional: Dr Franklin E. Rosales Científico Asociado, Traslado de Tecnología: Dr Luis Pocasangre Científico Asociado, Nematología: Thomas Moens C/o CATIE, Apdo 60 7071 Turrialba Costa Rica Fax : (506) 556 24 31 e-mail: [email protected] • Red Regional para Asia y el Pacífico Coordinator Regional: Dr Agustín B. Molina Científico Asociado, Traslado de Tecnología: Dra Inge Van Den Bergh C/o IRRI, Rm 31, GS Khush Hall Los Baños, Laguna 4031 Filipinas Fax : (63-49) 536 05 32 e-mail: [email protected] • Red Regional para Africa Occidental y Central Coordinator Regional: Dr Ekow Akyeampong Coordinator Regional de Información para África: Josué Tetang Tchinda Científico Asociado, Traslado de Tecnología: Kim Jacobsen C/o CRBP, B.P. 12438 Douala Camerún Fax: (237) 342 91 56 e-mail: [email protected] • Red Regional para África Oriental y del Sur Coordinator Regional: Dr Eldad Karamura Científico Asociado, Traslado de Tecnología: Dr Guy Blomme PO Box 24394 Kampala, Uganda Fax: (256-41) 28 69 49 e-mail: [email protected] • Centro de Tránsito INIBAP (ITC) Encargada: Ines Van Den Houwe Katholieke Universiteit Leuven Laboratory of Tropical Crop Improvement Kasteelpark Arenberg 13, B-3001 Leuven, Bélgica Fax: (32-16) 32 19 93 e-mail: [email protected]