File
Anuncio

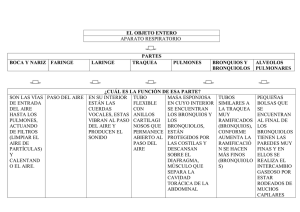
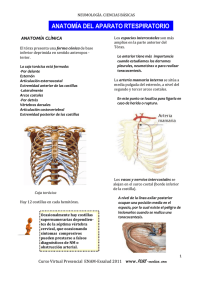
APARATO RESPIRATORIO Dra. Cynthia González Ruíz Profesor de carrera TC Patología General. I. FUNCIONES El aparato respiratorio cumple con importantes funciones metabólicas y de defensa: · Intercambio gaseoso · Regulación del equilibrio ácido-base · Regulación de la temperatura · Eliminación parcial o total de ciertas sustancias (siendo las más importantes: serotonina, norepinefrina, prostaglandinas, aldosterona, cortisol/cortisona, bradicinina) · Síntesis de prostaglandinas, leucotrienos; conversión de cortisona en cortisol y de angiotensina I en angiotensia II · Presencia de IgA en las vías respiratorias. Los macrófagos alveolares cumplen funciones inmunológicas y de fagocitosis · Reserva de sangre para compensaciones circulatorias De la respiración depende la captación de oxígeno del medio ambiente. El oxígeno es necesario para los procesos de oxidación del metabolismo intermedio que ocurren en los tejidos, con la producción de energía bajo la forma de ATP y, como productos residuales, CO2 y agua. De la respiración también depende la eliminación de CO2 formado, con lo cual interviene en la regulación del equilibrio ácido-base. La respiración sirve también para regular la temperatura corporal mediante la eliminación de calor bajo la forma de vapor de agua. La sangre es el vehículo que por intermedio del aparato cardiovascular, comunica los pulmones con los demás tejidos del organismo, siendo su objetivo final ceder a éstos el oxígeno transportado por la hemoglobina, con la participación reguladora del óxido nítrico contenido en los eritrocitos. El proceso de respiración tisular, está mediado por el sistema citocromo-oxidasa, en el que también interviene el óxido nítrico (que es un vasodilatador). El intercambio gaseoso se realiza fundamentalmente en los alvéolos pulmonares (Fig 1 y 2), pero también en los ductus alveolares y los bronquiolos terminales. Fig. 1 Figura 2. P1- Neumocito tipo I M- Macrófago alveolar P2- Neumocito tipo II La captación del oxígeno (hematosis) ocurre por difusión pasiva al existir un diferencial de tensión de oxígeno entre el aire y el medio interno a la altura de la membrana respiratoria. El oxígeno difunde a través de las células del epitelio alveolar (neumocitos tipo I-P1, Figura 2), que recubren el 95% de la superficie de los alvéolos) y del endotelio capilar, hacia la sangre; aquí es finalmente captado por la hemoglobina de los eritrocitos. Esencial para esta función es un flujo sanguíneo lento y a baja presión. La pared alveolar, con el grosor de una micra, se encuentra en íntima vecindad con un fino capilar, también de una micra de grosor. La luz de este capilar es de 7 a 10 micras de diámetro, lo que apenas permite el paso de uno a dos eritrocitos en fondo. Entre ambas membranas existe un espacio intersticial virtual, por lo que alvéolo y capilar constituyen una unidad funcional, que denominamos membrana respiratoria y / o Barrera hemato-aérea (Fig 3. Y Fig. 4). Por consiguiente, de la integridad de la misma depende la función respiratoria normal. Barrera hemato-aérea Figura 3 Figura 4 Un componente adicional muy importante es la presencia en los alvéolos de un complejo sistema surfactante lipo-proteico que recubre su superficie con una fina película protectora, la cual reduce la tensión superficial de los alvéolos y evita que éstos colapsen. Constituyen, además, parte del sistema de inmunidad innata. Este surfactante comienza a ser sintetizado hacia finales de la gestación por los neumocitos tipo II (Fig. 2) del epitelio pulmonar, bajo regulación de glucocorticoides. BRONQUIOS Figura 5. Representación esquemática de las unidades respiratorias de los pulmones. Los bronquios se forman por la bifurcación de la tráquea, uno derecho y otro izquierdo (Fig. 5), que se dirigen hacia el hilio del pulmón correspondiente, penetra en el pulmón y se ramifica en numerosas ramas cada vez de menor calibre, llamados bronquíolos, hasta llegar a los alvéolos pulmonares. En el bovino y el cerdo la tráquea se divide en tres bronquios: derecho, izquierdo y apical del pulmón derecho, que se desprende de la tráquea antes de los dos bronquios principales a la altura de la tercera costilla o espacio intercostal. En cada pulmón los bronquios primarios se dividen en bronquios secundarios. El número de bronquios secundarios coincide en humanos con el número de lóbulos que posee cada pulmón, dos el izquierdo y tres el derecho. La estructura de los bronquios (Fig. 6, 9 y 10) es similar a la de la tráquea, pero a medida que se van ramificando, va desapareciendo el cartílago progresivamente: de semilunas se convierten en placas dispersas (Figura 11). Además, el músculo liso es más patente y llega a ser una capa más visible que separa a la mucosa de la submucosa. El epitelio, que es pseudoestratificado cilíndrico ciliado en los bronquios de mayor calibre, al igual que en la tráquea, se convierte en pseudoestratificado cúbico, y con un espesor cada vez menor. Cuando el cartílago, las glándulas mucosas y las células caliciformes desaparecen y el músculo liso se hace también más escaso, aunque aún visible, los conductos entonces tienen un diámetro de aproximadamente 1 mm. A estos conductos se les denomina bronquiolos (Fig. 6, panel B y C). Figura 6. Órgano: Pulmón Especie: Rata. (Ratus norvegicus). Técnica: Secciones de parafina teñidas con tricrómico de Van Gieson. A. Bronquio B. Bronquiolo terminal C. Bronquiolo respiratorio D. Alveolo Figura 7. Aspecto de un pulmón de rata. La mayor parte del tejido está formado por alveolos, aunque también se aprecian bronquiolos terminales, bronquiolos y vasos sanguíneos. En este caso no se observan los glóbulos rojos porque el animal está perfundido. Los broquiolos (Fig. 6, panel B y C) no presentan cartílago, ni glándulas y, aunque los de mayor calibre tienen un epitelio pseudoestratificado cilíndrico ciliado, a medida que van disminuyendo su calibre, el epitelio se transforma en simple cilíndrico y posteriormente simple cúbico. Las glándulas mucosas también van disminuyendo su densidad a medida que disminuye el diámetro de los broquiolos. En todos ellos existe una capa de músculo liso bien patente. Los bronquiolos de mayor calibre se denominan terminales (Fig. 6, panel B) y los menor calibre son los bronquiolos respiratorios, los cuales tienen expansiones en su pared muy delgadas que forman unas oquedades denominadas alvéolos pulmonares (Fig. 6, panel D y Figura 7). Figura 8. Panel A Panel B Figura 8. Imágenes tomadas con el microscopio electrónico de barrido. La mayoría del tejido está formada por alveolos. En la imagen de la derecha se observa a mayores aumentos un bronquiolo terminal en la parte en que comunica con las cavidades alveolares. Los alvéolos (Fig. 8, panel A) son los lugares donde se produce el intercambio de gases entre la sangre y el aire. Son cavidades revestidas de un epitelio simple plano que aumentan enormemente la superficie de contacto con el aire. Cada alveolo está rodeado por un entramado capilar, de modo que la distancia entre los glóbulos rojos y el aire es muy escasa, favoreciendo el intercambio de gases. En humanos se estima que hay entre 150 y 250 millones de alveolos. A veces varios alveolos forman los conductos alveolares, que son expansiones de los broquiolos respiratorios (Fig. 8, panel B) formadas por numerosos alveolos asociados. Al final de los broquiolos respiratorios se suelen formar las lagunas alveolares, espacios abiertos a donde se abren numerosos alveolos. Figura 9. Figura 11. Figura 10. PULMONES Figura 12. Pulmones de los animales domésticos En los animales domésticos los pulmones tienen forma cónica, con la base en contacto con la cara anterior del diafragma y el vértice cerca de la entrada del tórax. El hilio de cada pulmón se encuentra en su parte medial, más o menos a la altura del lóbulo medio, y es el lugar por donde penetran el bronquio, la arteria pulmonar y los nervios pulmonares (autónomos) y además salen los vasos linfáticos y las venas pulmonares. En este lugar, la pleura mediastínica se refleja hacia el pulmón para convertirse en pleura visceral. Los pulmones se dividen en lóbulos, por medio de fisuras profundas que se inician en la parte ventral. En el canino, bovino y cerdo, el pulmón se divide en los lóbulos: craneal, medio, caudal y accesorio. En algunas especies el lóbulo craneal a su vez se subdivide en dos porciones, una craneal y otra caudal, por medio de una fisura poco profunda. En el equino, los pulmones no son lobulados, excepto por la presencia de un lóbulo intermedio en el pulmón derecho. La porción lateral de cada pulmón está en contacto con la pared lateral del tórax, menos en la escotadura cardiaca en donde el corazón alcanza dicha pared. PLEURA Es la membrana serosa que recubre la cavidad torácica y en parte los órganos contenidos dentro de ella. La pleura consta de dos sacos que se reflejan sobre cada pulmón. La pleura que se pega a las paredes de la cavidad torácica se llama pleura parietal y la que cubre los pulmones, pleura visceral. La unión de los dos sacos cerca de la línea media del tórax forma una doble capa de pleura que se conoce en este sitio con el nombre de pleura mediastínica, y el espacio que dejan se conoce con el nombre de espacio mediastínico, ocupado por varios órganos. Existen otras dos membranas serosas en el organismo: el pericardio, que recubre el corazón, y el peritoneo, que reviste la cavidad abdominal. II. LA RESPIRACIÓN La respiración es controlada por los centros respiratorios ubicados en el bulbo raquídeo y el puente troncoencefálico y por receptores en los cuerpos carotídeos. Estas estructuras son sensibles a variaciones en el pH del plasma y del líquido cefalorraquídeo (LCR), así como a la tensión parcial del O2 y del CO2 de la sangre; en menor grado intervienen también neuro-receptores pulmonares y de la pared torácica (Figura 13). La ventilación depende del volumen de aire que ingresa al pulmón; lo que en gran medida depende del trabajo de los músculos torácicos, a su vez controlados por el centro respiratorio. Figura 13 Como cualquier otra membrana mucosa que está en contacto directo con el medio ambiente, el aparato respiratorio tiene su propia flora bacteriana. Pero la flora respiratoria está restringida únicamente a la porción proximal del sistema de conducción (cavidad nasal, nasofaringe y laringe) mientras que las porciones distales del sistema de conducción (tráquea y bronquios), el sistema de transición (bronquiolos) y el sistema de intercambio (alvéolos) son membranas esencialmente estériles 26-28 . El sistema respiratorio está en constantemente relación por partículas (microorganismos, polvo, fibras), gases tóxicos (SO2, NO2, H2S, ozono) y vapores (amoniaco, formaldehído, acetona, gasolina). En condiciones normales, los gases inhalados son destoxificados, las toxinas son neutralizadas; las partículas son atrapadas y eliminadas, y los microorganismos son atrapados y eliminados. Cada región anatómica-histológica del aparato respiratorio tiene su propio mecanismo de defensa. En otras palabras, el sistema de conducción (de nariz a bronquios), sistema de transición (bronquiolos) y sistema de intercambio (alvéolos) tiene cada uno un mecanismo diferente de defensa. • Sistema de Conducción (Conchas nasales, tráquea y bronquios). El aparato mucociliar es el principal mecanismo de defensa del sistema de conducción, que incluye desde las fosas nasales hasta los bronquios (Figura 16). Todas estas estructuras están recubiertas por el llamado aparato mucociliar (Figura 11). Este aparato está formado por el epitelio pseudo-estratificado ciliar y las secreciones de las células caliciformes (moco). Cada célula ciliar del sistema de conducción tiene alrededor de 250 cilios los cuales producen alrededor de mil pulsaciones por minuto (1, 000/min) con un movimiento longitudinal promedio de 20 mm por min. Existen mecanismos auxiliares que facilitan el atrapamiento de partículas, vapores y gases por el aparato mucociliar (Figura 14). Por ejemplo, la generación de turbulencias de aire dentro de la cavidad nasal hace que las partículas mayores de 10 m sean atrapadas en el moco que recubre las conchas (cornetes) nasales. Las partículas de tamaño entre 3-10 m son atrapadas principalmente en las bifurcaciones de los bronquios en donde se originan fuerzas centrifugas en al aire inspirado al cambiar su dirección súbitamente. En síntesis, las partículas suspendidas en el aire de un tamaño de 3-10 m son atrapadas en el moco del aparato mucociliar (deposición) y de aquí son rápidamente eliminadas por el movimiento del moco hacia la faringe en donde son finalmente deglutidas. La IgA es la inmunoglobulina más abundante en el moco y una de sus principales funciones es inhibir la adherencia de patógenos a las células ciliadas. Figura 14. • Sistema de transición (bronquiolos – Figura 15). Sólo aquellas partículas de tamaño menor a las dos micras (<2 ) logran penetrar hasta los bronquiolos y alvéolos. En estas regiones profundas del pulmón, las partículas pequeñas se depositan en la membrana respiratoria mediante sedimentación o movimiento browniano. Los mecanismos de defensa de los bronquiolos son una combinación de los encontrados en los sistemas de conducción y de intercambio, más secreciones locales principalmente producidas por las llamadas células Clara. Figura 15. Bronquiolos Figura 16. Sistema de intercambio (alvéolos, Figura 17 y 18). Los alvéolos carecen de cilios y moco por lo que esta región pulmonar tiene un mecanismo de defensa especializado para protegerse de las partículas y patógenos inhalados. El principal mecanismo de defensa en el alveolo lo constituyen los macrófagos alveolares. Estas células fagocíticas se originan en la médula ósea de donde pasan a la sangre como monocitos sanguíneos, para después llegar al pulmón en donde permanecen un tiempo de “maduración” en el intersticio pulmonar. Durante el tránsito en el intersticio pulmonar adquieren la capacidad de fagocitar en un medio aeróbico. El número de macrófagos alveolares es proporcional al número de partículas respirables que llegan al pulmón; o sea que en un animal expuesto a un número más alto de partículas tendrá mayor número de macrófagos en los alvéolos. Las IgG e IgM son las inmunoglobulinas preponderantes en las secreciones alveolares y estos anticuerpos juegan un papel importante en la opsonización y fagocitosis por macrófagos alveolares. Las secreciones alveolares, particularmente el surfactante producido por los neumocitos tipo II, también contienen sustancias que favorecen la fagocitosis y actúan como antioxidantes para prevenir el daño celular causado por el estrés oxidativo. Figura 17. Figura 18. En conjunto, los mecanismos de defensa del aparato respiratorio son extremadamente eficientes en atrapar, destruir, eliminar, detoxificar agentes patógenos y gases tóxicos. Sin embargo, cuando estos mecanismos de defensa son deprimidos, las bacterias inhaladas colonizan fácilmente el pulmón causando infecciones respiratorias. De la misma manera, cuando gases, vapores o partículas tóxicas y agentes oxidantes sobrepasan los mecanismos de defensa, las células pulmonares son fácilmente dañadas causando considerables alteraciones pulmonares. Entre los factores externos más frecuentes que inhiben los mecanismos de defensa figuran las infecciones virales, el edema pulmonar, uremia, amoniaco, deshidratación, temperatura, humedad y estrés entre otros muchos. III. Mecanismos de resistencia Inmune del Aparato Respiratorio a la infección. La actividad del aparato mucociliar reduce la habilidad de los microbios de adherirse y proliferar en los pasajes aéreos del tracto respiratorio. La secreción de mucina y fluidos serosos en esta áreas contienen factores que inhiben la actividad bacteriana. Estos incluyen: lactoferrina, lizosima, proteínas surfactantes, fosfolipasa A2, inhibidor de leucoproteasa secretora, peroxidasa, serprocidinas, catelicidinas, defensinas y péptidos aniónicos. Estas substancias se ven incrementadas aparentemente en células epiteliales de bronquios y bronquiolos en respuesta a productos de bacterias Gram negativas. La inmunidad innata es importante para prevenir las infecciones microbianas en el tracto respiratorio, poco después del nacimiento, cuando las mucosas son inicialmente expuestas a una gran variedad de agentes infecciosos BIBLIOGRAFÍA 1. Jubb, Kennedy Palmer´s. 1993. Pathology of Domestic Animals. Academic th Press, 5 edition. Edited by M. Grant Maxie, DVM, PhD, DipACVP., Vol.2 2. McGavin, D. M., Carlton, W. W., Zachary, F. J., Thomson, G, R., 2001. Thomson’s Special Veterinary Pathology. Edited by St. Louis., London, Mosby. 3nd ed. 3. Banks, W. J. Applied Veterinary Histology. Second ed. Baltimore, Williams Wilkins., 1986. 4. Ackermann, R. M. And Brogden, A. K., 2000. Response of the ruminant respiratory tract to Mannheimia (Pasteurella) haemolytica. Microbes and Infection. 2: 1079-1088 5. Abbas, K. A., Lichtman, H. A. And Pober, S. J., 1999. Inmunología Celular y Molecular. Tercera edición. Interamericana- McGraw- Hill. Madrid; España. 6. Thomson’s Special Veterinary Pathology, Mosby, 2000