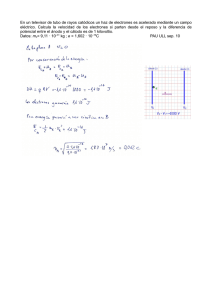
Célula Vegetal Pared Celular • limita al protoplasto y lo protege de posibles roturas provocadas por aumentos de volumen de la vacuola. • desempeña funciones importantes en la absorción, transporte y secreción de sustancias. • determina la forma de la célula. • suministra apoyo mecánico a las células. • media las interacciones de las células entre si. • actúa como barrera para el paso de moléculas grandes, pero permite el paso de moléculas pequeñas y iones. Pared Celular Composición • formada principalmente por moléculas de celulosa (polímeros lineares de glucosa). • las moléculas de celulosa se organizan en microfibrillas de 10 a 25 nm de diámetro. • cada microfibrilla se compone de un haz de 40 a 60 moléculas de celulosa orientadas en forma paralela y unidas entre sí por puentes de hidrógeno. • las microfibrillas se unen y giran sobre si mismas para formar macrofibrillas de 0,5 µm de ancho. Pared Celular Composición •el esqueleto celulósico de la pared está impregnado por una matriz entrelazada de moléculas no celulósicas. •algunas de éstas son los polisacáridos denominados hemicelulosa y otras son sustancias pécticas o pectinas, relacionadas químicamente con las hemicelulosas. •Otro constituyente en muchos tipos de células es la lignina, que sirve para dar rigidez a la pared y se encuentra células que tienen una función mecánica o de soporte. •También puede haber cutina, suberina y ceras, que son sustancias grasas que se encuentran comúnmente en las paredes de los tejidos externos de protección de la planta. •La cutina se encuentra en la epidermis y la suberina en un tejido secundario de protección, el súber, pero ambas se combinan con ceras y reducen la pérdida de agua en la planta. Pared Celular Capas de la Pared Celular Pared Celular Laminilla Media • Esta compuesta mayoritariamente por sustancias pécticas. • En tejidos leñosos generalmente la laminilla media está lignificada. • En los tejidos adultos es difícil de identificar porque se vuelve muy tenue. • Las paredes primarias y la laminilla media entre dos células contiguas se observan con microscopio óptico como una unidad que se denomina lámina media compuesta. • Cuando una pared celular se lignifica, el proceso comienza en la lámina media y se extiende posteriormente a la pared primaria y finalmente a la pared secundaria. Pared Celular Pared Primaria • Es la capa celulósica depositada en la pared antes y durante el crecimiento celular. • Además de celulosa, hemicelulosas y pectina, la pared primaria contiene glicoproteína y también se lignifica. • Las pectinas dotan de propiedades plásticas a la pared y le permite ser sometida a estiramiento. • Las células que se dividen continuamente contienen sólo paredes primarias. • Habitualmente no poseen un grosor uniforme y tienen áreas finas que se llaman campos primarios de las puntuaciones. Pared Celular Pared Secundaria • Se desarrolla luego que la célula ha detenido su crecimiento y la pared primaria terminó su elongación. • La pared secundaria es estructuralmente distinta de la primaria. • Son importantes en células especializadas que realizan funciones tales como el fortalecimiento y la conducción de agua. • Contiene mayor cantidad de celulosa que en las paredes primarias, pero carecen de sustancias pécticas y glicoproteínas. • La pared secundaria, además, es rígida y difícilmente deformable. • La matriz de la pared secundaría está compuesta por hemicelulosa. Pared Celular Pared Secundaria • A veces se pueden distinguir tres capas distintas llamadas S1, S2 y S3 para las capas externa, media e interna, respectivamente. • Las capas difieren entre sí por la orientación de las microfibrillas celulósicas y se encuentran en determinadas células del xilema o leño. • La estructura laminada de estas paredes secundarias aumenta la resistencia de la pared. • Las microfibrillas celulósicas están adosadas más densamente que sobre la pared primaria. Pared Celular Crecimiento en Espesor de la Pared Celular • Los materiales de la fase amorfa (sustancias pécticas, hemicelulosas y proteínas) son transportados por vesículas de los dictiosomas. • La celulosa es producida por complejos de enzimas celulosa-sintetasas que están incluidas en la membrana plasmática en forma de rosetas. • Las rosetas de enzimas reciben las moléculas de glucosa del lado citoplasmático de la membrana y las unen formando fibras de celulosa que se depositan en el lado externo. Pared Celular Crecimiento en Espesor de la Pared Celular • A medida que las rosetas se desplazan en la membrana, crecen las microfibrillas, que se depositan al azar y quedan entrelazadas. • Cuando se deposita la pared secundaria, las rosetas se agregan en grupos casi hexagonales, que tienen hasta 16 filas de rosetas dispuestas geométricamente. • El grupo se mueve en la membrana plasmática como una unidad, depositando filas paralelas de microfibrillas. Pared Celular Crecimiento en Espesor de la Pared Celular Pared primaria Pared secundaria Pared Celular Crecimiento en Extensión de la Pared Celular • En células isodiamétricas las microfibrillas se depositan formando una red irregular y la pared aparece como una sucesión de redes de microfibrillas (teoría de la red múltiple). • En células alargadas se depositan perpendicularmente al eje de crecimiento de la célula, pero las microfibrillas muestran cambios en su orientación. • Se pasa de una orientación casi horizontal a otra casi vertical cuando terminó el alargamiento y al mismo tiempo se depositan por dentro otras capas con diferente orientación. Pared Celular Comunicaciones Intercelulares Plasmodesmos Campo primario de puntuación Puntuación simple Puntuaciones • Puntuación simple ramificada Puntuación areolada • sin toro • con toro Perforaciones Plasmodesmos • Son canales que atraviesan la pared celular uniendo los citoplasmas de células adyacentes y facilitando la comunicación intercelular. • Están agrupados en zonas adelgazadas de las paredes primarias, constituyendo un campo primario de puntuacion o puntuación primordial. B Campo de puntuaciones primarias con plasmodesmos A Pared primaria Célula A Pared primaria Laminilla media A B Célula B Plasmodesmos Diagrama de plasmodesmo entre dos células Campo primario de puntuación en vista superficial (MEB) • El modelo actual sugiere que existe un tubo membranoso comprimido derivado del RE que se encuentra en el centro del canal y recibe el nombre de desmotúbulo. • Existen proteínas globulares asociadas a proteínas integrales que se proyectan como radios desde el desmotúbulo hacia la membrana plasmática dividiendo el cilindro citoplásmico y formando microcanales Plasmodesmos Los plasmodesmos pueden presentarse en tres estados: Cerrados: se caracterizan por falta de intercambio de moléculas entre células vecinas y tal estado puede ser transitorio o permanente. Abierto: se caracteriza por el libre movimiento de iones y metabolitos. Dilatados: son una extensión de los plasmodesmos abiertos y permiten el movimiento de macromoléculas. • En paso entre los diferentes estados se debe al complejo actina/miosina que se encuentra dispuesto helicoidalmente a la largo del desmotúbulo conectando la MP con el mismo. • Este complejo actúa regulando el tamaño del anillo citoplásmico y de los microcanales. • Concentraciones elevadas de Ca producen el cierre transitorio de los plasmodesmos. Puntuaciones • Puntuación simple Pared secundaria A B CORTE ÓPTICO Puntuaciòn simple Puntuación simple Cèlula A Célula B VISTA SUPERFICIAL Par de puntuaciones simples Entre las cèlulas A y B Abertura externa PARTES DE LA PUNTUACIÓN Canal Membrana de cierre Abertura interna • la pared secundaria se interrumpe Pared primaria abruptamente Célula A • se presenta en células parenquimáticas, fibras y esclereidas Pared primaria Célula B Laminilla media Puntuaciones • Par de puntuaciones simples Lumen Pared secundaria Canal Fibras del floema Fibra 1 Fibra 2 Membrana de cierre • Puntuación simple ramificada • Se forman debido a la fusión de los canales de dos o más puntuaciones vecinas, debido al incremento en grosor de la pared celular. Braquiesclereidas en Pyrus communis, pera • Puntuación areolada sin toro • la pared secundaria al depositarse, hace un reborde o aréola formando la cámara de la puntuación que se abre al lumen celular a través de la abertura de la puntuación. VISTA SUPERFICIAL CORTE OPTICO Pared secundaria Abertura Abertura interna externa Areola Abertura externa Abertura interna Càmara de la puntuaciòn Membrana de cierre Pared primaria Cèlula A Laminilla media Pared primaria Cèlula B • Puntuación areolada sin toro • El canal puede tener forma de embudo aplanado, y entonces las aberturas interna y externa difieren: la interna es lenticular o lineal y la externa es pequeña y circular. • En un par de puntuaciones, las aberturas internas están frecuentemente dispuestas en cruz, en relación con la disposición inclinada de las fibrillas de celulosa en la pared secundaria. • Puntuación areolada con toro • las puntuaciones areoladas presentan un engrosamiento central secundario lignificado. • la zona periférica llamada margo está formada sólo por fibrillas dispuestas radialmente, ya que la matriz y la laminilla media han desaparecido por acción enzimática. VISTA SUPERFICIAL CORTE ÓPTICO Pared secundaria Abertura Abertura interna externa Aréola Abertura externa Abertura interna toro Cámara de la puntuación Margo Pared primaria Célula A Laminilla media Pared primaria Célula B • Puntuación areolada con toro VISTA SUPERFICIAL Abertura Abertura interna externa toro Margo • Puntuación areolada con toro • En los elementos conductores, el paso del agua de una célula a otra se realiza a través de la puntuación y el toro interviene en la regulación de dicho mecanismo. • Cuando el toro está en posición media, el agua pasa libremente a través del margo; cuando el toro se une a una de las aberturas impide el paso del líquido. Puntuaciones areoladas con toro en traqueidas de Pinus silvestris Perforaciones • son otro tipo de comunicación intercelular, en el que hay una interrupción de la pared primaria y laminilla media, además de la pared secundaria. • Se presenta en células de los tejidos de conducción, en los vasos del xilema, donde forman las placas de perforación. Vasos del xilema con perforaciones Pared Celular Comunicaciones Intercelulares Plasmodesmos Campo primario de puntuación simple Puntuaciones • sin toro areolada Perforaciones • con toro Sustancias ergásticas • Su nombre proviene del griego "ergon” que significa trabajo. • es decir que son productos del metabolismo celular, de reserva o de desecho, que se acumulan en la pared celular, en las vacuolas o en plastos. Almidón • Es el carbohidrato de reserva de las plantas superiores y se acumula en los amiloplastos. • Es el alimento básico más importante de la humanidad. • Se halla en células parenquimáticas de corteza, médula y tejidos vasculares de tallos y raíces. • Se obtiene almidón comercial de Manihot esculenta y Solanum tuberosum. 10x Granos de almidón en tubérculo de Ipomoea batatas (Convolvulaceae), batata. Cristales • Se forman generalmente en las vacuolas, y se los considera como productos de excreción, aunque que en ciertos casos son reutilizados. • El componente más común es el Oxalato de Ca, que resulta de la acumulación intracelular de Ca en los tejidos. • Puede tener diferentes formas agujas = rafidios columnas = estiloides prisma = prismáticos estrella = drusas arena = arenilla cristalina Cristales de oxalato de calcio Cristal prismático Drusas Rafidios Cristales Cristales de carbonato de calcio 40x Cistolitos en hoja de Ficus elastica, gomero Cistolito • Los cristales de carbonato de Ca no son comunes en plantas superiores. • Generalmente están asociados con las paredes celulares formando una estructura llamada cistolito, que posee un pedúnculo celulósico. • En ciertas células de la epidermis de monocotiledóneas llamadas células silícicas, se forman cuerpos de sílice de forma característica: en las gramíneas son amorfos, generalmente no angulares. Proteínas Aleurona en cariopse de trigo • Se almacenan en el citoplasma dentro de vacuolas o plastos, bajo la forma de cristaloides. • Las que se almacenan en las vacuolas son sintetizadas en el RE rugoso, de donde pasan a los dictiosomas y luego a grandes vacuolas de proteína de reserva. • Las vacuolas se convierten en un cuerpo proteico sólido o grano de aleurona en el tejido de reserva maduro (cotiledones de semillas de Leguminosas y capa de aleurona del cariopse de las Gramíneas). • Cada grano de aleurona está limitado por el tonoplasto y puede tener una matriz amorfa o una matriz que incluye un cristaloide proteico y un globoide amorfo no proteico (Ricinus). Taninos • Son un grupo heterogéneo de derivados fenólicos muy frecuentes en el cuerpo vegetal. • Aparecen en las vacuolas como gránulos finos o gruesos, o como cuerpos de formas variadas, de color amarillo, rojo o marrón, o pueden impregnar las paredes. • Abundan en hojas, tejidos vasculares, peridermis, frutos inmaduros y cubiertas seminales. • Pueden estar en células especiales, los idioblastos tánicos. • Sirven como defensa contra el ataque de parásitos (hongos y microorganismos) o la acción de animales fitófagos. Plastos Plastos Cloroplastos y gerontoplastos Etioplastos con pigmentos sin pigmentos globulosos fibrilares o Cromoplastos tubulosos cristalinos membranosos Leucoplastos proplastos amiloplastos proteinoplastos oleoplastos Transformaciones de los plastos Plastos Cromoplastos Solanum lycopersicum Cromoplastos de color rojo en células de pimiento • Almacenan pigmentos como carotina (amarillo o anaranjado), licopina (rojo) y xantofila (amarillento). • Se encuentran en pétalos u otras partes coloreadas tales como frutos (tomate, pimiento) y raíces (zanahoria). • Los cromoplastos se desarrollan a partir de cloroplastos por síntesis y acumulación de pigmentos carotenoides acompañada por la desaparición del sistema de tilacoides. • La diferenciación de un cromoplasto no es un fenómeno irreversible; los cromoplastos pueden diferenciarse en cloroplastos perdiendo los pigmentos y desarrollando tilacoides. Plastos Cromoplastos Hay cuatro categorías de cromoplastos según su estructura: 1) globulosos: los pigmentos se acumulan en gotas junto con lípidos. 2) fibrilares o tubulosos: los pigmentos se asocian con fibrillas proteicas. 3) cristalinos: los pigmentos se depositan como cristaloides asociados con las membranas tilacoides. 4) membranosos: membranas arrolladas helicoidalmente. Cromoplastos según su estructura Plastos Amiloplastos • Almacenan almidón. • Tienen forma muy variada: esféricos, ovales o alargados. • Normalmente muestran una deposición en capas alrededor de un punto llamado hilo. • El hilo puede ser céntrico o excéntrico. • Cuando hay más de un hilo se forman granos compuestos. Granos de almidón fotografiados con MEB Estructura de los amiloplastos Oryza sativa, arroz Zea mays, maíz Plastos Proplastos • Son pequeños plastos indiferenciados, incoloros que están en células meristemáticas de las raíces y tallos. • Son precursores de otros plastos muy diferenciados como cloroplastos, cromoplastos o amiloplastos. • Si se detiene su desarrollo por ausencia de luz, puede formar cuerpos prolamelares, formados por membranas tubulares. Los plastos que tienen cuerpos prolamelares se denominan etioplastos. Etapas de transformación de un etioplasto en cloroplasto Plastos Cloroplastos son los organelos encargados de realizar la fotosíntesis tienen forma alargada y miden 510 µm x 2-4 µm número variable (20-40) una doble membrana, separadas por un espacio intermembranoso la membrana externa presenta porinas la membrana interna, es prácticamente impermeable Plastos Cloroplastos la membrana interna esta organizada en sacos aplanados llamados tilacoides se agrupan en estructuras llamadas granas (pilas de monedas) tiene pocos fosfolípidos y muchos glucolípidos neutros la matriz se denomina estroma ribosomas 70S y una molécula de ADN circular son organelos autónomos capaces de autoduplicarse Cloroplastos Genoma del Cloroplasto • Oscila entre 120 y 160 kb y contiene alrededor de 120 genes. Cloroplastos Cloroplastos Internalización de Proteínas • Las proteínas son dirigidas por secuencias Nterminales de 30 a 100 aminoácidos llamadas péptidos de tránsito. • Estas dirigen el transporte a través de las dos membranas del cloroplasto y que después son eliminadas mediante escisión proteolítica. • Los péptidos son reconocidos por el complejo de translocación de la membrana externa del cloroplasto (complejo Toc). • Luego las proteínas se transfieren al complejo de translocación interno (Tic) y son transportadas al estroma. • Como en las mitocondrias, para la internalización se requieren chaperonas moleculares y energía en forma de ATP. • Sin embargo, a diferencia de las mitocondrias, los péptidos de tránsito no tienen carga positiva. • Por ello, el transporte de proteínas al interior del cloroplasto no requiere un potencial eléctrico a través de la membrana. Cloroplastos Transporte de Proteínas al Tilacoide • Primero son internalizadas en el estroma, como ya se ha descrito. • Luego son marcadas y dirigidas para atravesar la membrana del tilacoide por una segunda secuencia señal hidrofóbica, que queda expuesta tras la lisis del péptido de tránsito. • La secuencia señal hidrofóbica dirige la translocación del polipéptido a través de la membrana del tilacoide. • Esta secuencia finalmente es eliminada en la luz por una segunda escisión proteolítica. Fotosíntesis • Durante la fotosíntesis se utiliza la luz solar para dirigir la síntesis de glucosa a partir de CO2 y H2O. • La fotosíntesis se realiza en dos fases distintas: • En la fase lumínica, la energía de la luz solar dirige la síntesis de ATP y NADPH, acoplada a la formación de O2 a partir del H2O. • En la fase oscura (no requiere luz solar), el ATP y el NADPH producido en la fase lumínica dirigen la síntesis de glucosa. • En las células eucariotas, tanto la fase lumínica como la fase oscura de la fotosíntesis tienen lugar en los cloroplastos. • Las reacciones de la fase lumínica ocurren en la membrana tilacoidal y las reacciones de la fase oscura en el estroma. Fotosíntesis Flujo de electrones a través de los fotosistemas I y II • La luz solar es absorbida por los pigmentos fotosintéticos. • La absorción de la luz excita un electrón a un estado energético más elevado, convirtiendo la energía de la luz solar en energía química. • Los pigmentos están organizados en fotocentros en la membrana del tilacoide, cada uno de los cuales contiene cientos de moléculas de pigmento. • Los pigmentos en cada fotocentro actúan como antenas, absorbiendo la luz y transfiriendo la energía de sus electrones excitados a una molécula de clorofila que sirve como centro de reacción. • La clorofila del centro de reacción transfiere su electrón de alta energía a una molécula aceptora de una cadena de transporte de electrones. • Los electrones de alta energía se transfieren entonces a través de una serie de transportadores de membrana, acoplados a la síntesis de ATP y de NADPH. Fotosíntesis • Las proteínas que intervienen en la fase lumínica de la fotosíntesis se organizan en cinco complejos en la membrana del tilacoide. • Dos de ellos son fotosistemas, que absorben luz y se transfieren a las clorofilas del centro de reacción. • Los electrones de alta energía se transfieren a través de una serie de transportadores tanto en los fotosistemas como en un tercer complejo de proteínas, el complejo citrocromo bf. • Al igual que en las mitocondrias, la transferencia de electrones está acoplada a la transferencia de protones a la luz del tilacoide, estableciéndose así un gradiente de protones a través de la membrana. • La energía almacenada se obtiene mediante un cuarto complejo, la ATP sintetasa, que acopla el flujo de protones a través de la membrana con la síntesis de ATP. Fotosíntesis • En la fase oscura, el ATP y el NADPH producidos se utilizan para sintetizar carbohidratos a partir del CO2 y H2O. • En total, el ciclo consume 18 moléculas de ATP y 12 de NADPH por cada molécula de glucosa sintetizada. Fotosíntesis • Deben pasar 24 electrones por la cadena de transporte de electrones para generar suficiente NADPH para sintetizar una molécula de glucosa. • Estos electrones se obtienen de la conversión de 12 moléculas de H2O a seis moléculas de O2, lo que produce la formación de seis moléculas de O2 por cada molécula de glucosa. • No está claro, sin embargo, si el paso de estos mismos 24 electrones por la cadena de transporte de electrones es también suficiente para generar los 18 ATP que se requieren para el ciclo de Calvin. Fotosíntesis Flujo Cíclico de Electrones • Es una segunda ruta de transporte de electrones que genera ATP sin la síntesis de NADPH. • La energía de la luz obtenida en el fotosistema I se utiliza para la síntesis de ATP en lugar de para la síntesis de NADPH. • Los electrones ricos en energía son transferidos del fotosistema I al complejo citocromo bf. • La transferencia de electrones del citocromo bf se acopla al transporte de protones a través del tilacoide. • La plastocianina devuelve luego estos electrones al fotosistema I en un estado energético más bajo, completando un ciclo de transporte de electrones. • Por lo tanto, la transferencia de electrones desde el fotosistema I puede generar ATP o NADPH, dependiendo de las necesidades metabólicas de la célula. Fotosíntesis Síntesis de ATP • Por cada par de electrones transportados en el fotosistema II se transportan dos protones al interior del tilacoide y de dos a cuatro protones al complejo citocromo bf. • Como se necesitan cuatro protones para la síntesis de una molécula de ATP, el paso de cada par de electrones a través de los fotosistemas rinde entre 1 y 1,5 moléculas de ATP. • El flujo cíclico de electrones tiene un rendimiento inferior, entre 0,5 y 1 moléculas de ATP por cada par de electrones.
 0
0
Anuncio







Descargar
Anuncio
Añadir este documento a la recogida (s)
Puede agregar este documento a su colección de estudio (s)
Iniciar sesión Disponible sólo para usuarios autorizadosAñadir a este documento guardado
Puede agregar este documento a su lista guardada
Iniciar sesión Disponible sólo para usuarios autorizados