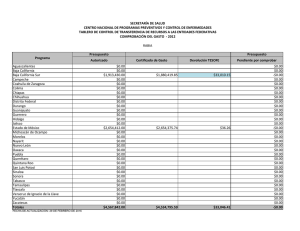
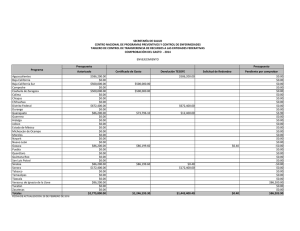
Pinos San Pedro-Preliminares.p65
Anuncio

Pinos San Pedro-Preliminares.p65 2 04/02/2005, 05:04 p.m. EL BOSQUE DE CONÍFERAS DE LA SIERRA DE SAN PEDRO MÁRTIR, BAJA CALIFORNIA, MÉXICO Pinos San Pedro-Preliminares.p65 3 04/02/2005, 05:04 p.m. Pinos San Pedro-Preliminares.p65 4 04/02/2005, 05:04 p.m. EL BOSQUE DE CONÍFERAS DE LA SIERRA DE SAN PEDRO MÁRTIR, BAJA CALIFORNIA, MÉXICO José Delgadillo Rodríguez Secretaría de Medio Ambiente y Recursos Naturales Instituto Nacional de Ecología Pinos San Pedro-Preliminares.p65 5 04/02/2005, 05:04 p.m. Pinos San Pedro-Preliminares.p65 6 04/02/2005, 05:04 p.m. Contenido Prólogo 7 Introducción 8 Capítulo 1. Generalidades Primeros habitantes Primeros exploradores La misión La actividad ganadera La actividad forestal Estatus de conservación Otras propuestas 9 9 9 10 11 13 15 16 Capítulo 2. Caracterización física Fisiografía Geología Edafología Hidrología 17 17 21 24 25 Capítulo 3. Bioclimatología Clima mediterráneo Bioclimatología Diagramas climáticos Diagrama de balance hídrico Pisos bioclimáticos Piso mesomediterráneo Piso supramediterráneo 26 27 28 35 35 44 49 50 Pinos San Pedro-Preliminares.p65 7 04/02/2005, 05:04 p.m. Capítulo 4. Fitogeografía Región californiana Provincia martirense 48 57 57 Capítulo 5. Composición florística Flora Formas biológicas Endemismo Consideraciones florísticas especiales 59 59 60 64 66 Capítulo 6. Descripción de la vegetación Bosque de pinus Bosque de pinus de montaña Bosque de Pinus quadrifolia y Pinus monophylla Bosque de Pinus jeffreyi Bosque mixto Bosque de Pinus contorta var. murrayana Bosque de Juniperus Bosque de Cupressus Vegetación riparia Praderas 70 72 73 75 79 81 86 87 89 91 95 Capítulo 7. Comunidades fitosociológicas Generalidades Bosque de pino piñonero y juniperus Bosques de pinus de montaña symphoricarpoioreophlii-Pinetum jeffreyi Potentillo rimicolae-pinetum murrayanae 95 95 99 106 107 Bibliografía 109 Glosario 121 Anexo I. Catálogo florístico 127 Anexo II. Catálogo fotográfico 143 Pinos San Pedro-Preliminares.p65 8 04/02/2005, 05:04 p.m. Figuras Figura 1. Ubicación de la Sierra de San Pedro Mártir Figura 2. Corte transversal de la Sierra de San Pedro Mártir Figura 3. Leyendas de un diagrama climático Figura 4. Diagrama climático de la estación meteorológica No. 95 Figura 5. Diagrama Balance hídrico de la estación meteorológica No. 95 Figura 6. Pisos de vegetación de Baja California Figura 7. Distribución aproximada de los pisos bioclimáticos y de vegetación de la Sierra de San Pedro Mártir Figura 8. Fitogeografía de Baja California Figura 9. Representación esquemática del bosque de coníferas 18 21 38 38 42 47 48 56 74 Fotos Foto 1. Vertiente oeste de la Sierra de San Pedro Mártir Foto 2. Vertiente este de la Sierra de San Pedro Mártir Foto 3. Vista del Picacho del Diablo (3100 m), desde el área de Observatorio mayor Foto 4. Exposición de roca granítica en la Sierra de San Pedro Mártir Foto 5. Una tormenta de verano vista desde la vertiente oeste de la Sierra de San Pedro Mártir Foto 6. Vista invernal (con nieve) de la zona de Vallecitos Foto 7. Piso bioclimático mesomediterráneo Foto 8. Piso bioclimático supramediterráneo Foto 9. Comunidades de Pinus quadrifolia FOTO 10. Comunidades de Pinus quadrifolia Foto 11. Panorámica hacia el oeste del bosque de coníferas Foto 12. Pinus jeffreyi y Quercus agrifolia a lo largo de cañadas Foto 13. Comunidad de Populus tremuloides (álamo) 19 19 20 19 23 34 51 53 76 77 81 81 94 Gráficas Grafica 1. Distribución estacional de la precipitación Pinos San Pedro-Preliminares.p65 9 32 04/02/2005, 05:04 p.m. Cuadros Cuadro 1. Temperatura promedio mensual de dos estaciones meteorológicas Cuadro 2. Datos de temperatura de la estación meteorologica No. 95 Cuadro 3. Valores de precipitación media anual Cuadro 4. Valores reportados de precipitación en diferentes periodos Cuadro 5. Datos de evaporación (ET) de la estación meteorológica No. 95 Cuadro 6. Las diez familias con el mayor numero de taxa Cuadro 7. Géneros con el mayor número de taxa y su forma biológica Cuadro 8. Porcentaje de las formas biológicas de la flora vascular Cuadro 9. Algunos de los taxa predominantes en el bosque de coníferas Cuadro 10. Relación de los taxa endémicos registrados Cuadro 11. Índice de abundancia-cobertura de Braun-Blanquet Pinos San Pedro-Preliminares.p65 10 04/02/2005, 05:04 p.m. 30 30 32 33 43 60 61 62 63 65 96 Prólogo Este trabajo reúne información sobre los recursos naturales, particularmente la vegetación, de la Sierra de San Pedro Mártir en Baja California. Conjunta datos provenientes de importantes estudios desarrollados en la zona así como información obtenida por el autor a lo largo de más de 22 años como profesor de botánica de la Facultad de Ciencias de la Universidad Autónoma de Baja California. Su propósito es describir estos bosques borelaes únicos en México, considerados una isla biogeográfica en la península de Baja California, y su relación con el entorno físico (geología, edafología y clima). A partir de colectas y datos de herbarios se hace un análisis de la flora registrada en esta sierra y se describe la riqueza florística del área. Se presenta un listado de las especies de plantas que encontramos en la región y un catálogo ilustrado de las especies de plantas representativas del área y que pueden considerarse como las características de la estructura del bosque. Este trabajo representa un aporte para conocer mejor los aspectos físicos y bióticos del estado de Baja California, y de manera especial la riqueza natural de una porción muy importante no solo de esta entidad sino de nuestro país. La Sierra de San Pedro Mártir cuenta con una larga historia de decretos de protección para sus recursos naturales (reserva forestal, parque nacional y reserva forestal nacional), por lo que el contenido de este libro puede ser de utilidad para la elaboración del programa de manejo de esta área natural protegida. Prólogo Pinos San Pedro-A Prólogo.p65 11 04/02/2005, 05:06 p.m. 11 12 El bosque de coníferas Pinos San Pedro-A Prólogo.p65 12 04/02/2005, 05:06 p.m. Introducción La Sierra de San Pedro Mártir es un sistema de alta montaña localizado en el estado de Baja California. Geológicamente formado por granodioritas, es la continuación meridional de la cordillera montañosa californiana. La vegetación en sus partes altas presenta un bosque de coníferas de carácter templado seco, similar en su fisonomía y composición florística general a los bosques del sur de California (EE.UU.). Sus características climáticas de tipo mediterráneo, únicas en México, favorecen la condición relictual de este tipo de bosque boreal en la península de Baja California. Desde la perspectiva de su bioclima, presenta dos pisos de vegetación, el supramediterráneo y mesomediterráneo superior; por su diversidad florística y sus comunidades vegetales ha sido considerado una isla biogeográfica y una provincia fitogeográfica, la Martirense, diferente de la Californiania. El bosque de coníferas se distribuye a partir de los 1,500 msnm, hasta prácticamente los 3,100 m en el Picacho del Diablo, el pico más alto de la península. Su estructura es muy abierta, con un sotobosque muy pobre. Sus praderas tienen una alta diversidad de herbáceas perennes, y representan zonas de alta capacidad de captación de agua, principalmente durante el invierno. Este bosque de coníferas presenta un alto grado de conservación (90%), sin duda debido a lo alejado de los centros urbanos y lo poco accesible durante la época de invierno. Cuenta con los siguientes decretos: reserva forestal (1923), parque nacional (1947), reserva forestal nacional (1951). En octubre de 1996, y posteriormente en 2003, se firmó el convenio entre la SEMARNAP, la Comisión Nacional de Áreas Naturales Protegidas (CONANP) y el Introducción Pinos San Pedro-B Introducción.p65 13 04/02/2005, 05:07 p.m. 13 Gobierno del Estado de Baja California para la transferencia de acciones de manejo y conservación de esa zona, también clasificada como una región terrestre prioritaria para la Conservación por la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO) y otras instituciones. 14 El bosque de coníferas Pinos San Pedro-B Introducción.p65 14 04/02/2005, 05:07 p.m. Capítulo 1. Generalidades Los primeros habitantes Los kiliwas, etnia del grupo lingüístico yumano (Meigs 1935, 1939, en Minnich et al. 1994) son los habitantes más antiguos de la Sierra de San Pedro Mártir, y de cuya presencia durante miles de años dan cuenta los vestigios arqueológicos de la región. La Sierra es, y ha sido para este grupo, su casa y su fuente de alimento (Estrada 1991). Hinton (1957 en Minnich et al. 1994) señaló que la mayoría de los kiliwas se asentaban a lo largo de Arroyo de León, una área que consideraban como su «reserva» y que les fue otorgada por el gobierno. Fueron cazadores nómadas y recolectores de plantas, utilizando mucho los recursos de la Sierra, pero también practicaron la agricultura (Meigs 1939, Owen 1963 en Minnich et al. 1994). El principal producto obtenido de estas actividades era el piñón en otoño, que colectaban hasta hace una década, cuando fue quemado su piñonal; por su parte, los paipai, otro grupo indígena de la región, siguen practicando su recolección hasta la fecha (Estrada 1997 en Minnich y Franco-Vizcaíno 1999). Primeros exploradores La Sierra de San Pedro Mártir fue observada a distancia por Ulloa (1539) y después por los padres jesuitas Eusebio Kino y Juan María Salvatierra (1701), durante los viajes marítimos en el Golfo de California. Posteriormente fue explotada, con algunos asentamientos durante un período relativamente corto, en la época turbulenta de la historia de las misiones, a finales del siglo XVIII (Meigs 1935 en Minnich et al. 1994). Generalidades 15 El primer europeo en visitar la región fue el jesuita Wenceslaus Linck, ministro de la misión de San Borja, en el centro de Baja California (Burrus 1966, Meigs 1935 en Minnich et al. 1994). En 1776 Linck cruzó el sudeste de la Sierra de San Pedro Mártir y exploró las planicies desérticas al oriente de la Sierra. Después de su expulsión en 1768, los jesuitas fueron reemplazados en la región por la orden franciscana que organizó la histórica expedición de 1769 a través del noroeste de Baja California hasta San Diego, atravesando el margen occidental de la Sierra. Este viaje fue parte de un esfuerzo por resistir la expansión de Rusia, que extendía sus asentamientos desde Alaska hacia el sur, sobre la costa del Pacífico (Bolton 1927, Meigs 1935 en Minnich et al. 1994). Finalmente, tras el concordato firmado en 1772 por el virrey y los franciscanos y dominicos, los primeros tomaron a su cargo la Alta California y los segundos el sistema de misiones de la Baja California. La misión La misión dominica de San Pedro Mártir de Verona fue una de las misiones más aisladas y originales de las establecidas en el Nuevo Mundo. Aunque tuvo una vida relativamente breve (1794-1806), este puesto avanzado de la orden de los dominicos se distinguió como la única misión dedicada principalmente a la ganadería y no a la agricultura, como era la norma. Su terreno cultivable fue de menor extensión que el de otras misiones; sin embargo, un sistema de riego extensivo proporcionaba cosechas adecuadas. Los dominicos establecieron la misión en una «pradera con agua rodeada por bosque» llamada Casilepe, en el límite norte de la pradera La Grulla. Casilepe resultó ser inadecuada por la severidad de su clima. Situada en el valle a 2,100 msnm, fue la misión establecida a mayor altitud, tanto de la Alta como de la Baja California (Foster 1991). Poco tiempo después, los dominicos seleccionaron finalmente un lugar más templado localizado a una menor altitud (1550 m) a lo largo del arroyo El Horno, 10 km al este de San Isidoro. La misión cerró en 1806 (Foster 1992 en Minnich et al. 1994), y actualmente se considera una las misiones californianas más remotas. El sitio de la misión consiste en ruinas de adobe, objetos dispersos en una extensa superficie, algunos canales y campos anteriormente cultivados (Foster 1991). 16 El bosque de coníferas La actividad ganadera En la Sierra de San Pedro Mártir, y principalmente su zona boscosa, se han realizado actividades de pastoreo desde la época de los misioneros dominicos (Meling 1991). Arrillaga, uno de los primeros exploradores (1796), reportó que la misión enviaba ganado a pastar al valle de La Encantada durante el verano, y a otras áreas de mejor calidad (Meigs 1935 en Minnich et al.1994). Entre 1828 y 1915 la actividad ganadera era importante, como lo demuestran los registros de la marca de herrar de los ganaderos de los poblados de San Telmo y Santo Domingo, origen de los actuales ganaderos de la región, siendo imposible ser ganadero sin contar con el pastoreo en la Sierra de San Pedro Mártir (Meling 1991). En 1915 se formaron las compañías borregueras, mismas que posteriormente empezaron a incursionar en la Sierra. Este hecho molestó a los ganaderos de bovino quienes consideraban injusto que personas ajenas a la región vinieran a invadir lo que no era de ellos, además de que el hábito de manejo y pastoreo de la ganadería ovina y bovina era diferente. Este último pastorea en forma dispersa, come pasto sin perjudicar la raíz, y no molesta al pino nuevo, mientras que los ovinos pastorean en rebaños de aproximadamente 2,000 cabezas comiendo pasto, raíces, follaje y pinos nuevos; al terminarse el alimento se movían a otro lugar dejando completamente sobre-pastoreada la zona (Meling 1991). Desde 1924 y hasta la década de 1960, la Sierra fue sometida a un pastoreo irracional autorizado de ganado lanar con un promedio de 6,000 cabezas. Según Meling (1991), en 1963 o 1964 se negó la subida de la borregada al parque nacional y con su salida se inició la recuperación del bosque y sus pastizales, dando un nuevo auge a la ganadería bovina, actividad que hasta la fecha se sigue desarrollando, casi de la misma manera que antaño. La temporada óptima de subida al parque nacional es de fines de mayo a mediados de junio, dependiendo qué tanta lluvia haya caído en el año, y permanecen hasta finales de octubre, al iniciar el frío. La ganadería de los poblados de San Telmo y Santo Domingo depende en un 50% a 75% del pastoreo en la Sierra (Meling 1991). Los principales pastizales en la Sierra son La Grulla, La Encantada, Santo Tomás y Vallecitos. De acuerdo con Iturbe et al. (1984) los ganaderos han llevado a pastar un número muy grande de animales (se calcula que suben alrededor de Generalidades 17 5,000 cabezas), de abril a octubre de cada año, sin tomar en cuenta las condiciones climáticas ni el estado de recuperación de los pastos. Excepto en la zona sur del parque, en los parajes conocidos como La Grulla y La Encantada, donde supuestos propietarios han realizado la división del valle en potreros, es donde se observa un mejor manejo del pasto, lo que no ocurre en el resto del parque donde, por el contrario, se observa una degradación paulatina con el avance de especies indeseables (Iturbe et al. 1984). Según la Comisión Técnica Consultiva para la Determinación Regional de los Coeficientes de Agostadero (COTECOCA), el coeficiente de agostadero para el Parque Nacional Sierra de San Pedro Mártir es un máximo de 1865 “unidades animal” que pueden pastar. Esta misma Comisión refiere que las condiciones edafológicas, climáticas y de pastoreo, impiden que la cobertura vegetal potencial, la cual puede ser hasta 90%, no se en la mayor parte de la región superior al 30%. Las actividades ganaderas en el ejido El Bramadero están bajo el control de 20 o 30 propietarios y 150 a 200 familias dependen de esta actividad (Meling 1995). El ejido cubre un área de aproximadamente 300,000 hectáreas, la mayor parte sobre la ladera occidental de la sierra. La actividad forestal En 1945 se estableció la explotación en diferentes zonas, para lo cual se realizaron estudios sobre 65,000 ha y a partir del 10 junio de 1946, por acuerdo presidencial, se faculta a la Secretaría de Agricultura y Ganadería para autorizar aprovechamientos de madera y resina; así fue otorgada una concesión (27 de noviembre de 1946) a la Compañía de San Pedro Mártir S. de R.L. para aprovechar 153, 328 m3 de madera en rollo durante diez años; también se le otorga concesión a la Compañía Manuel Ortega Bojorques, S. de R.L. (sin fecha disponible). Mediante el decreto del 21 de febrero de 1947, publicado en el Diario Oficial de la Federación el 26 de abril del mismo año, la zona concesionada fue declarada parque nacional, posteriormente, el 27 de julio de 1951 se declara reserva forestal nacional a la Sierra de San Pedro Mártir y se establece una en ella veda definitiva e indefinida de servicio de montes, por lo que al oponerse al interés público se cancelan de pleno derecho las concesiones de explotación forestal otorgadas a las compañías antes referidas (SEMARNAP 2000). 18 El bosque de coníferas De acuerdo con el Inventario Forestal del Estado de Baja California de 1968 (SARH 1977), las superficie estudiada en las regiones de la Sierra de Juárez y la Sierra de San Pedro Mártir abarcarón 605, 317 ha correspondiendo el 81% (491, 479 ha) a superficie forestal; 396, 807 ha, los chaparrales y 86, 550 ha coníferas. Del área total de coníferas, el 60 % correspondió a bosques comerciales accesibles, siendo la Sierra de San Pedro Mártir la de mayor superficie de este tipo de bosques, tanto en valor absoluto como relativo. La Sierra de Juárez es el macizo forestal más extenso del estado con 342, 113 ha y la de San Pedro Mártir cuenta con 149, 366 ha, siendo el 80% de arbolado comercial. Esto explica que, no obstante ser menor la superficie de coníferas en la región, y en comparación con Sierra de Juárez, su volumen fue 2.7 veces mayor. De acuerdo con Salas (1997), en las primeras exploraciones realizadas en la Sierra de San Pedro Mártir en busca del mejor sitio para la instalación de un observatorio astronómico, se encontraron cabañas con sierras, a manera de aserraderos, que se utilizaban para la corta y aprovechamiento de la madera de pinos. Según Minnich et al. (1994) se observaron camiones de carga transportando largas tablas de madera de pino Jeffreyi camino abajo del observatorio. También Minnich et al. (1997) refieren que en junio de 1996 se observó a un grupo americano de taladores con equipo pesado para transportar troncos. En 1995 se le otorgó al ejido El Bramadero un permiso de aprovechamiento por cinco años para saneamiento forestal con cortas selectivas para la extracción de arbolado afectado por plagas y enfermedades, equivalentes a 43,478 m³ en rollo (40,240 m³ de Pinus jeffreyi y 3,238 m³ de Abies concolor) (Sierra 1995). De acuerdo con Minnich y Franco-Vizcaíno (1999) el periódico La Jornada publicó que se había otorgado a Diamond Mountain Resources, de Almanor, California, una concesión para explotar los recursos biológicos de la Sierra de San Pedro Mártir. La actividad extractiva otorgada al ejido El Bramadero fue suspendida por las autoridades correspondientes ya que, de acuerdo a la Comisión Nacional de Áreas Naturales Protegidas, hubo corta de arbolado sano dentro del parque nacional. También el Registro Agrario Nacional (RAN) determinó en septiembre del 2003, que el aprovechamiento referido se encuentraba dentro del parque nacional. Generalidades 19 Estatus de conservación En la Sierra de San Pedro Mártir se encuentra un bosque de coníferas único en México, y que es continuación de este tipo de vegetación presente también en California (Delgadillo 1998). Esta zona boscosa esta considerada para su protección dentro las siguientes categorías: I Reserva forestal. Decreto presidencial del 4 de octubre de 1923. I Parque nacional. Decreto presidencial del 26 de abril de 1947. I Reserva forestal nacional. Decreto presidencial del 25 abril de 1951, pu- blicado el 27 de julio del mismo año, confirmando el decreto anterior del 4 de octubre de 1923. I Área de interés público para su conservación y restauración de su riqueza forestal. Decreto del 17 de febrero de 1975. El 11 de julio de 1947 se delegó al gobierno del Territorio Norte de Baja California la administración del parque nacional, con facultades para efectuar cortes de saneamiento y aclareo, así como de reforestación. Estas facultades fueron derogadas por el Acuerdo del 3 de marzo de 1948 (Hernández et al. 1977). Por decreto presidencial del 11 de octubre de 1978 el parque nacional se transfiere para su administración a la Secretaría de Asentamientos Humanos y Obras Públicas (SAHOPE) del Gobierno del Estado de Baja California, una vez que la Secretaría de Agricultura y Recursos Hidráulicos (SARH), hubiera realizado su limpieza y saneamiento del parque nacional. Al desaparecer la SAHOPE, la administración del parque nacional quedó bajo la jurisdicción de la Secretaría del Medio Ambiente, Recursos Naturales y Pesca (SEMARNAP ). Dentro del parque nacional y reserva forestal hay un área concesionada a la Universidad Nacional Autónoma de México (UNAM) para la instalación y operación de un observatorio astronómico (Gobierno del Estado de Baja California, SFA y SEMARNAP 1996). En octubre de 1996 se firmó un convenio de concertación entre el Ejecutivo federal, por conducto de la SEMARNAP, el Gobierno del Estado de Baja California, la Universidad Nacional Autónoma de México, la Universidad Autónoma de Baja California, el Instituto Nacional de Ecología y el Ayuntamiento de Ensenada, con el propósito de constituir el Comité Técnico del 20 El bosque de coníferas Parque Nacional Sierra de San Pedro Mártir. Su el objeto, la restauración, conservación, desarrollo y vigilancia de dicha área natural protegida (Gobierno del Estado de Baja California, SFA y SEMARNAP 1996). El 29 de octubre de 2003 se firmó el convenio de modificación al acuerdo de coordinación, siendo la Secretaría de Fomento Agropecuario, del Gobierno del Estado de Baja California, el organismo responsable en la administración de esta zona. Otras propuestas I Reserva de la Biosfera de la Sierra de San Pedro Mártir, bajo el programa MAB-México y MAB-UNESCO (Barry 1991, Minnich et al. 1994, Minnich 1995). I Área piloto de conservación dentro del Programa de Áreas Protegidas 1995-2000 (SEMARNAP 1995). I Región prioritaria para la conservación de la Sierra de San Pedro Mártir, bajo el programa de Áreas Protegidas de la Comisión para el Estudio de la Biodiversidad en México (CONABIO/ PRONATURA/ WWF/ FMCN/ USAID/ TNC/ INE 1998). Generalidades 21 Capítulo 2. Caracterización física Fisiografía La Sierra de San Pedro Mártir se encuentra en la porción norte de Baja California, fisiográficamente está formada por montañas, laderas y valles centrales (figura 1). El sistema montañoso recorre prácticamente toda la península, con pendientes a menudo muy abruptas que descienden directamente al Mar de Cortés y mucho más suaves hacia el océano Pacífico. Así, la vertiente oriental de la Sierra es muy accidentada y en distancias relativamente cortas (30 km), se desciende desde los aproxima-damente 3,100 m en Picacho del Diablo hasta los 600 m en la Laguna del Diablo y las grandes planicies del desierto de San Felipe, hasta llegar al Golfo de California. Esta vertiente se caracteriza por sus grandes cañones y paredes (algunos de ellos de difícil acceso), así como algunos arroyos perennes (por ejemplo. La Providencia, El Diablo, La Esperanza) (Delgadillo 1999) (fotos 1 y 2). La Sierra de San Pedro Mártir es la más alta de varios terrenos batolitos que comprenden las cordilleras peninsulares y del sur de California; limita hacia el norte con la falla de Agua Blanca y hacia el este con una gran falla escarpada de más de 2,500 m de longitud, que está separada por las cuencas de Valle Chico y Valle San Felipe. Hamilton (1971 en O’Connors y Chase 1989) refirió que esta característica es una de las más imponentes de Norte América. Las mesetas de la cumbre, las laderas hacia el sur y oeste, y las superficies inclinadas son cortadas por una o dos fallas asociadas con aproximadamente 1,000 m de relieve (Woodford y Harris 1938 en O’Connors y Chase 1989). Las altitudes máximas son La Corona de Arriba (2,565 m), BaCaracterización física 23 jío de las Viejas (2,590 m), Cerro Botella Azul (2,800 m), Área del Observatorio (2,840 m) y el Picacho del Diablo (3,100 m) (foto 3). FIGURA 1. UBICACIÓN DE LA SIERRA DE SAN PEDRO MÁRTIR, BAJA CALIFORNIA Sierra de San Pedro Mártir Una alta meseta de más de 1,800 metros se extiende por más de 15 km en dirección este-oeste y recorre más de 50 kilómetros de longitud en dirección NO-SE, uniéndose en el norte con el Valle de la Trinidad y Sierra de Juárez. Continúa hacia el sur hasta la abandonada Misión de San Pedro Mártir, a 1,600 m. La figura 2 muestra un perfil hipotético transversal de la Sierra de San Pedro Mártir (Álvarez 1983). La amplia presencia de sedimentos fluviales del Terciario y la extensa erosión de la superficie de las mesetas altas de la cordillera peninsular indican que el desarrollo fue esencialmente terminado en el Eoceno medio. Por encima de los 1,500 m existe un sistema de drenaje hacia el oeste, mientras que entre los 1,000 y 1,500 m los drenajes son más uniformes hacia el 24 El bosque de coníferas FOTO 1. VERTIENTE OCCIDENTAL DE LA SIERRA DE SAN PEDRO MÁRTIR FOTO 2. VERTIENTE ORIENTAL DE LA SIERRA DE SAN PEDRO MÁRTIR Caracterización física 25 FOTO 3. VISTA DEL PICACHO DEL DIABLO (3,100 M), DESDE EL ÁREA DEL OBSERVATORIO MAYOR FIGURA 2. CORTE TRANSVERSAL DE LA SIERRA DE SAN PEDRO MÁRTIR A TRAVÉS DE LOS PUNTOS A, DONDE SE ENCUENTRA EL OBSERVATORIO ASTRONÓMICO Y EL PUNTO B, LLAMADO “LA CORONA”, EL PUNTO MÁS ALTO EN EL LADO OESTE DE LA SIERRA. EL EJE HORIZONTAL ESTÁ MARCADO EN KILÓMETROS DESDE EL OBSERVATORIO. EL EJE VERTICAL EN METROS A PARTIR DEL NIVEL DEL MAR (ÁLVAREZ 1983). A Altitud (m) 3,000 B 2,000 Sierra de San Felipe 1,000 0 63.1 26 50 El bosque de coníferas 25 0 Distancia (km) 25 50 62.4 sudoeste (O’Connors y Chase 1989). En general, el sistema de drenaje de numerosos arroyos es mucho más somero hacia el oeste y muy abrupto hacia el este (Delgadillo 1999). Geología La Sierra de San Pedro Mártir, al igual que Sierra Nevada, la forman rocas batolíticas del Mesozoico y paredes de rocas metasedimentarias; y aparentemente el levantamiento del bloque se produjo durante el Cenozoico tardío (Chase y Wallace 1986 y 1988 en O’Connors y Chase 1989). El levantamiento de Sierra Nevada está asociado con la formación de la cuenca regional y la extensión de la cordillera (Hamilton y Myers 1966 en O’Connors y Chase 1989), mientras la Sierra de San Pedro Mártir y el Picacho del Diablo están más relacionadas con los cambios de los límites entre las placas del Pacífico y Norte América en los últimos 14 millones de años (O’Connors y Chase 1989). Las evidencias indican que el levantamiento de la Sierra de San Pedro Mártir empezó después del Mioceno temprano y que el mayor levantamiento probablemente ocurrió durante el Plioceno y Cuaternario, asociado con la separación de la península de Baja California de Norte América hacia las placas del Pacífico. Un posterior levantamiento de la parte alta de la sierra ocurrió en el Cuaternario al menos entre los últimos 5.5-3.5 millones de años (O’Connors y Chase 1989). Las rocas ígneas, que comprenden la mayoría de las áreas con crestas de la cordillera peninsular, incluida la Sierra de San Pedro Mártir, son fundamentalmente tonalitas y granodioritas (foto 4) (Silver y Chapell 1988 en O’Connors y Chase 1989). Las partes más altas de la Sierra están subrayadas por una larga intrusión de granodioritas con rocas metasedimentarias (Gastil et al. 1975, Gastil 1991, O’Connors y Chase 1989) y complejos batolíticos al este de la cordillera peninsular, los cuales fueron emplazados hace 105-80 millones de años (Silver y Chapell 1988, en O’Connors y Chase 1989). Desde Vallecitos hasta las praderas de La Grulla y La Encantada, la sierra está constituida por un plutón masivo de granito, que da lugar a cerros de topografía irregular. Los cuerpos graníticos más resistentes a la intemperización forman superficies de roca fracturada espectaculares en Cerro Venado Blanco. Hacia el sur, desde el Cerro Botella Azul hasta La Grulla y La Encantada, hay grandes exposiciones de rocas metamórficas que se Caracterización física 27 intemperizan formando laderas de topografía más homogénea. Las unidades metamórficas afloran en el centro-norte de Vallecitos, cerca del Observatorio Astronómico Nacional, así como en los cerros del extremo norte de la sierra (Minnich y Franco-Vizcaíno 1997b). La proporción de exposición, así como el carácter generalmente reciente de las rocas de esta área, son ideales para una variedad de estudios geoquímicos, geofísicos y tectónicos. En el norte de la región central de la sierra se encuentra expuesto un gigantesco y heterogéneo plutón (gran intrusión de magma) de 400 km² (Gastil 1991). Edafología De acuerdo al mapa de suelos del estado de Baja California (INEGI 2001), la mayor parte de la sierra cuenta con suelos de tipo Litosol con textura media y en su ladera oeste por Xerosoles de textura gruesa y media. Los primeros son suelos presentes en todos los climas y sustentan muy diversos tipos de vegetación; se caracterizan por tener una profundidad menor de 10 cm hasta la roca, tepetate o caliche puro y de textura media. Los segundos son suelos típicos de las zonas áridas y semiáridas del centro y norte de México, siendo su vegetación natural matorrales y pastizales con capas superficiales de color claro y muy pobre en humus; debajo de ellas puede haber un subsuelo rico en arcillas, o bien muy semejante a la capa superficial. Estos suelos son de textura franco-arenosa o arenosa-franca, de color gris oscuro a gris-café oscuro, moderadamente profundos (60-80 cm), bien drenados y muestran un perfil poco desarrollado (xeropsamments típicos) (INEGI 2001). En las praderas, los suelos son más profundos (más de 150 cm), y parecen estar influenciados por un fuerte gradiente de humedad desde las partes bajas hacia las zonas ligeramente más altas; un desarrollo moderado con zonas moteadas (Haplaquepts Aéricos), que parece deberse a un ciclo anual de condiciones aeróbicas durante el invierno y primavera cuando los suelos están saturados, seguido de condiciones aeróbicas en el periodo seco de verano y otoño (INEGI 2001). En la mayor parte de la zona boscosa los suelos son de origen granítico (Delgadillo 1998), seguidos en extensión por los formados a partir de esquistos (Minnich et al. 1994); la roca madre está situada a 30 o 40 cm de profundidad (Passini et al. 1989). Estos suelos graníticos son moderadamente ácidos (Fran- 28 El bosque de coníferas FOTO 4. EXPOSICIÓN DE ROCA GRANÍTICA EN SIERRA DE SAN PEDRO MÁRTIR co-Vizcaíno et al. 1992 en Minnich et al. 1994). Los horizontes superficiales son ligeramente más ácidos comparados con los de la Sierra de Juárez que oscilan de pH medio de 4.5 a 6 (Passini et al. 1989). Son suelos no salinos y pobres en nutrientes, con concentraciones de nitrógeno, fósforo, potasio y magnesio críticamente bajas y menores que las mínimas reportadas para suelos agrícolas en California (Franco-Vizcaíno et al. 1992 en Minnich et al. 1994). En áreas fuera del parque nacional y hacia la vertiente que desciende al océano Pacífico, se observan suelos perturbados con pinos y encinos y rocas de pizarras a 1,750 m. En exposición sur, aproximadamente a 1,500 m, comienzan suelos rojos y arcillosos, y al descender a 1,100 m los suelos tienen estructuras en bloques, angulares y prismáticos, con inclusiones de concreciones cálcicas entre las fisuras (Hernández et al. 1977). HIDROLOGÍA Los mayores caudales hidrológicos se encuentran en las partes más altas de la vertiente oeste, y desembocan en el océano Pacífico. En cambio, en la Caracterización física 29 vertiente oriental la caída de los arroyos es mucho más pronunciada debido a la orografía y todos terminan en la llamada Laguna del Diablo, situada en el inicio de terrenos aluviales del desierto de San Felipe. Algunos arroyos occidentales llevan agua durante todo el año, aumentan su caudal en los periodos de lluvia invernal y aportan grandes cantidades de agua a partir de los deshielos. Los principales arroyos originados en las partes altas de la Sierra son: San Rafael, La Tasajera-El Potrero-Valladares, Santa Cruz, San Antonio, Los Pinos, El Picacho, Cieneguita de Soto, Meling y La Corona (Delgadillo 1999). 30 El bosque de coníferas Capítulo 3. Bioclimatología Generalidades García y Mociño (1968), mencionan que la ubicación del noroeste de México, en el extremo occidental de la gran masa continental de Norteamérica, tiene dos consecuencias notables que condicionan en gran medida su aridez. En primer lugar, está sometida a los efectos de la celda de alta presión del Pacífico durante la mayor parte del año; y en segundo término, la influencia de la corriente de California que discurre de norte a sur, paralela a la costa de la península de Baja California. La corriente oceánica fría de California, modifica y controla el régimen meteorológico general, sobre todo a bajas altitudes, aumentando la zona atmosférica que inhibe la formación de nubes de tipo convectivo en las cercanías de la costa del Pacífico (Álvarez 1983). De acuerdo al sistema de clasificación de Köppen (1948 en Rzedowski 1978) a la península de Baja California le corresponde la clasificación de clima tipo B, o seco, que cubre la mayor parte de México, sobre todo en la mitad septentrional de su territorio, y en particular en el altiplano, planicies costeras noroccidentales y nororientales. Los climas de categoría BW (seco desértico), predominan en la mayor parte de la península y en menor medida encontramos la categoría Bs (seco estepario). Los climas de tipo C, o templados y húmedos, son característicos de las zonas de montaña de México. La categoría Cs (con temporada lluviosa en época fría del año), es propia de las montañas de Baja California (Rzedowski 1978). Bioclimatología 31 Clima Mediterráneo El noroeste de la península de Baja California, incluida la Sierra de San Pedro Mártir presenta un clima clasificado como mediterráneo, semejante al existente en otras partes del mundo, como la propia cuenca mediterránea, centro de Chile, sur de África y sudoeste de Australia (Delgadillo 1998). Walter (1985) en función de la estacionalidad de temperaturas y precipitaciones (valores absolutos), variación altitudinal, y en ciertos casos la naturaleza del suelo, reconoció nueve biomas zonales o zonobiomas en el mundo: ecuatorial, tropical, subtropical árido, mediterráneo, templado cálido, templado, templado árido, boreal y ártico. La Sierra de San Pedro Mártir se encuentra dentro del Zonobioma IV, que corresponde al tipo Mediterráneo. El clima mediterráneo de esta región apareció a finales del Terciario (Axelrod 1975), por lo que la mayor parte de su flora nunca ha estado en contacto con las otras regiones dotadas de idéntico clima, lo que explicaría en parte el elevado grado de endemismo de esta región (Raven 1988). Dunn et al. (1976) refieren que estas áreas se caracterizan por un patrón distintivo donde se combinan lluvias moderadas en invierno y temperaturas frías debajo del óptimo para el crecimiento de las plantas, seguidas de veranos secos, calientes y sin lluvias, donde las temperaturas son favorables para la actividad de las plantas (foto 5). En nuestra región más del 75% de la precipitación se concentra en el periodo que va de octubre a abril, y con menos del 20% en los siguientes seis meses (Pase 1982). Particularmente en las montañas de la Sierra de San Pedro Mártir existe una región con gran proporción de tiempo despejado, siendo una de las tres regiones sin nubes en el mundo (Mendoza et al. 1972 en Álvarez 1983), lo que hizo óptima para la instalación en 1967 del Observatorio Astronómico Nacional. Bioclimatología La bioclimatología es la ciencia que estudia la relación existente entre los organismos vivos y el clima, y difiere de la climatología en que sus modelos y unidades descriptivas han sido delimitados con base en las especies y biocenosis (bioindicadores), que le otorgan su estabilidad, principalmente botánica hasta el presente (Rivas-Martínez 1987). El conocimiento de la di- 32 El bosque de coníferas FOTO 5. UNA TORMENTA DE VERANO EN LA SIERRA DE SAN PEDRO MÁRTIR VISTA DESDE LA VERTIENTE OESTE námica en la vegetación puede ser interpretada con los cambios climáticos (Dansereau 1957). Von Humboldt (1807), Schouw (1823), Meyen (1846) y De Cadandolle (1855 en Woodward 1987) pusierón énfasis en la importancia del clima en los patrones observados en la distribución de la vegetación del mundo. Los resultados naturales de estas relaciones provocaron la búsqueda de los mecanismos mediante los cuales el clima ejerce este control (Woodward 1987). Clima, fisiografía y geología juegan un papel central en la determinación de la distribución de las clases de suelos, tipos de vegetación, floras y faunas. Los biogeógrafos están de acuerdo que el clima es el primer factor en el control de esta distribución (Brouiller y Whetstone 1993 en Woodward 1987). La ausencia de equivalencias en la delimitación de zonas climáticas se debe al irregular énfasis de varios factores y elementos (Dansereau 1957). Trewartha (1942 en Dansereau 1957) sugirió como factores la latitud, la distribución de los continentes, el relieve, la presión barométrica, y las corrientes marinas; y como elementos el viento, la temperatura y la precipitación. Bioclimatología 33 Los factores determinan el comportamiento de los elementos, pero son estos últimos los que influyen en los organismos vivos. De entre los elementos climáticos, la precipitación y temperatura son determinantes fundamentales en la distribución de los ecosistemas en la Tierra (Dansereau 1957). Temperatura. La distribución de la temperatura media anual y su comparación con un mapa topográfico de la república mexicana indica de manera elocuente, que el principal factor determinante de este parámetro climático es la latitud y en un lugar secundario, la influencia altitudinal. El gradiente térmico, en función de la altitud, varía de una región a otra como consecuencia de factores diversos, los cuales pueden jugar un papel destacado: pendiente, altura relativa del macizo montañoso, humedad, latitud, etc. (Rzedowski 1978). Una situación extraordinaria se presenta en la vertiente occidental de casi toda la península de Baja California, donde, por efecto de la corriente fría de California, se establece una inversión térmica durante una parte del año y por consiguiente, al ir ascendiendo altitudinalmente, se comprueba que entre 0 y 800 m la temperatura media aumenta en vez de disminuir (García y Mociño 1968, Álvarez y Maisterrera1977; en Álvarez 1983); hacia el sur de la península y del Golfo de California no se presenta esta capa de inversión. La Sierra de San Pedro Mártir es parte de la provincia climática del noroeste (zona montañosa), Las estaciones metereológicas se encuentran en la parte alta de la Sierra de San Pedro Mártir (estación No. 95 y Observatorio), no cuentan con datos anuales constantes de por lo menos 20 años que nos permitan calcular promedios anuales tanto de temperatura como de precipitación (Delgadillo 1999). Sin embargo, con los datos disponibles de siete años de la estación metereológica No. 95 se obtuvo una temperatura anual promedio de 10.6 °C (Delgadillo 1995). Por su parte, Álvarez (1983) reportó para la zona del Observatorio (2,830 m) una temperatura promedio anual de 7.2 °C, para un periodo de seis años (1969-1974), siendo enero el mes mas frío con 1.5 °C en promedio y el mes de julio el más calido con 16.3 °C promedio. El cuadro 1 muestra datos comparativos de las dos anteriores, mientras que el cuadro 2 presenta los valores máximos, mínimos y absolutos de la estación No. 95 (Delgadillo 1999). 34 El bosque de coníferas CUADRO 1. TEMPERATURA PROMEDIO MENSUAL DE DOS ESTACIONES METEOROLÓGICAS EN LA SIERRA DE SAN PEDRO MÁRTIR: ÁREA DEL OBSERVATORIO (ÁLVAREZ 1983) Y ESTACIÓN NO. 95 (SARH) E F M A M J J A S O N D Anual Años Fuente 1.5 0.6 -1.0 4.6 9.6 13.7 16.3 14.3 13.6 8.0 3.0 2.1 7.2 1969-74 Álvarez 1983 4.1 5.4 5.9 8.2 10.6 16.3 18.2. 16.9 15.2 11.1 9.2 6.7 10.6 1984-91 Estac. No. 95 (SARH) Fuente: Delgadillo1999. CUADRO 2. DATOS DE TEMPERATURA DE LA ESTACIÓN METEREOLÓGICA NO. 95 (2,080 MSNM) PROMEDIOS Máxima absoluta del mes cálido (M’) Media de las máxima absoluta mes más cálido (M) Media máximas absolutas mes más frío (Mm) Mínimas absoluta del mes más frío (m’) Media mínima absolutas del mes más frío (m) Media mínima del mes más frío (Mminf) Media de las máxima (Mmax) Media de las máxima del mes más frío (Mmxmf) 35 °C 30 °C 15.4 °C - 14.5 °C - 5.8 °C 2.8 °C 25.6 °C 20.8 °C Precipitación. Los bosques de coníferas de la región californiana aparecen en un rango rango de precipitación es muy variable (Barbour 1988, Major y Taylor 1988, Hasting y Turner 1965). El noroeste de la península tiene un régimen regular de precipitación pluvial muy definido durante los meses de invierno (noviembre-abril) con escasa, lluvias el resto del año. La humedad durante la época invernal proviene del noroeste en forma de tormentas que originan fuertes lluvias y nieve, esta última a altitudes mayores a 1,100 m. Las tormentas tropicales y ciclones que se originan en el Bioclimatología 35 Pacífico, debido a procesos de convección, provocan lluvias durante el verano de (julio a septiembre) en las partes altas de las Sierras de Juárez y San Pedro Mártir, desiertos y algunas veces en la costa del Pacífico (Miranda et al. 1997, Delgadillo 1999). La gráfica 1 muestra los porcentajes de precipitación por cada periodo estacional: invierno 220.7 mm (43%), primavera 120.9 mm (23%), verano 96.1 mm (19%), y otoño 80.9 (16%). En el periodo invierno-primavera, que va desde el mes de diciembre hasta mayo, la precipitación media es de 341.6 mm (66%) (Delgadillo 1999). El cuadro 3 muestra los valores de la media anual de precipitaciones reportadas para la parte alta de Sierra de San Pedro Mártir, mientras que el cuadro 4 presenta valores de precipitación en diferentes periodos. La mayor precipitación promedio mensual se presenta durante los meses de marzo (106.6 mm) y enero (99.5 mm), mientras que junio tiene los valores más bajos (0.7 mm) (Delgadillo 1999). Nieve. Además del agua de lluvia es importante considerar el aporte extraordinario de humedad procedente por el deshielo de la nieve (Delgadillo 1995). Cuando el sistema de nubes alcanza las costas se inicia un proceso denominado “forzamiento orográfico”, que obliga a las masas de aire a elevarse; en consecuencia, mientras más inclinada esté la pen- GRÁFICA 1. DISTRIBUCIÓN ESTACIONAL DE LA PRECIPITACIÓN EN LA SIERRA DE SAN PEDRO MÁRTIR 50 Invierno Precipitación (mm) 40 30 Primavera 20 Verano Otoño 10 0 36 El bosque de coníferas Estaciones Fuente: Delgadillo 1999 CUADRO 3. VALORES DE PRECIPITACIÓN MEDIA ANUAL REPORTADOS EN LA SIERRA DE SAN PEDRO MÁRTIR PRECIPITACIÓN (MM) SITIO PERIODO 250-500 400 700 550 Observatorio (2830) 1969-1974 ——— ——— La Corona (2600 m) 1989-1993 Planicie central (2400 m) ——— 518 Estación No.95 (2080 m) 1984-1991 FUENTE Álvarez 1983 Miranda et al. 1997 Escoto-Rodríguez 1994 Minnich y FrancoVizcaíno 1997 Delgadillo 1995 Fuente: Delgadillo 1999. CUADRO 4. VALORES REPORTADOS DE PRECIPITACIÓN EN DIFERENTES PERIODOS EN LA SIERRA DE SAN PEDRO MÁRTIR PERIODO Diciembrefebrero Marzomayo Junioagosto Septiembrenoviembre Media invernal Total invernal PRECIPITACIÓN (MM) SITIO PERIODO FUENTE 220.7 Estación No.95 1984-1991 SARH 120.7 Estación No.95 1984-1991 SARH 96.1 Estación No.95 1984-1991 SARH 80.9 Estación No.95 1984-1991 SARH 450-550 400 Oeste y ——— centro de la planicie Ladera este ——— Total invernal 260-280 ——— Total invernal 550-560 ——— 1989-1990 (periodo seco) 1990-1992 (periodo húmedo) Minnich et al. 1997 Minnich et al. 1997 Minnich et al. 1997 Minnich et al. 1997 Fuente: Delgadillo 1999. Bioclimatología 37 diente del terreno y más altas estén las cimas de las montañas, más suben las masas de aire. Estas características atmosféricas en condiciones de tormenta puramente extratropical ocasionan nevadas en las cimas de las Sierras de Juárez y San Pedro Mártir (Reyes 1993). La nieve es causante, en buena parte, del mantenimiento de la elevada biomasa de estos bosques, aun en sus áreas más secas (Pase 1982). El porcentaje de nieve va desde 15%, a 1,700 m, hasta 80%, a los 2,600 m (Minnich 1986 en Minnich et al. 1997). En el bosque a baja altitud (1,980 m), la nieve se derrite rápidamente y se infiltra en el suelo produciendo un drenaje profundo y significativo. En cambio en el bosque de mayor altitud (2,470 m) la nieve permanece de dos a tres meses y el agua de deshielo aparentemente corre sobre el suelo congelado pero por debajo de la nieve. De esta manera, menos agua entra al perfil del suelo durante el invierno en las áreas de bosque alto, y en consecuencia, existe menor drenaje profundo y agua almacenada en el suelo que en los bosques bajos (Munich et al. 1994). Humedad relativa. Rzedowski (1978) menciona que este elemento climático es de gran interés ecológico porque a menudo juega un papel de FOTO 6. VISTA DE VALLECITOS DURANTE EL INVIERNO 38 El bosque de coníferas importancia en la distribución de la vegetación, sin embargo, el número de estaciones meteorológicas que registran el contenido de humedad es escaso. La proximidad al océano Pacífico, combinada con la altitud provoca la existencia de una alta humedad relativa en la Sierra de San Pedro Mártir, con el aporte de nieblas (criptoprecipitaciones), principalmente en laderas sombreadas de orientación norte y por encima de los 2, 400 m. Estas laderas se distinguen por presentar comunidades casi puras de Abies concolor, de donde “cuelgan” líquenes del género Ramallina (Delgadillo 1999). Heladas. Dunn et al. (1976) refieren que las áreas mediterráneas se caracterizan por un patrón distintivo, donde se combinan lluvias moderadas en invierno y temperaturas frías debajo del óptimo para el crecimiento de las plantas, seguido de veranos secos y calientes, sin lluvias intermedias, lo que da lugar a las temperaturas que resultan favorables para la actividad de las plantas. De acuerdo a Visher (1954 en Hanes 1965), las plantas en nuestra región mediterránea tienen de 250 a 360 días sin heladas. Los meses de heladas probables se establecen con la media de las mínimas mayores a cero grados centígrados, pero con las mínimas absolutas por debajo de los cero grados. Si la media de las mínimas es menor a cero grados, se dice que es un mes de heladas seguras.También, puede calcularse el periodo sin heladas. En todo caso, estos parámetros constituyen un buen indicador del termoclima, y de forma aproximada podemos hacer corresponder el periodo de heladas probables y seguras en distintos meses y pisos de vegetación (Delgadillo 1999). Periodo de actividad vegetal (PAV). Hace tiempo que los ecólogos se interesan por evaluar los datos de la temperatura en función de su efecto sobre la reproducción y crecimiento de las plantas. Los datos mensuales de temperatura y precipitaciones deben separarse, por lo menos estacionalmente, ya que tienen diferente significado fisiológico (Daubenmire 1957). El periodo de actividad vegetal se define como el conjunto de meses del año con medias mensuales iguales o mayores a 7.5 ° C (Álvarez 1992). Se observa con frecuencia que el periodo de actividad en las plantas es relativamente tardío o temprano, dependiendo del comienzo de las temperaturas adecuadas en la primavera y de que la actividad siempre comienza más tarde a latitudes y altitudes progresivamente más altas. En promedio, Bioclimatología 39 los eventos fenológicos (relaciones entre los factores climáticos y los fenómenos periódicos en los organismos) se presentan cuatro días más tarde con cada grado de latitud en dirección al polo, cinco grados de longitud en dirección este (en Norteamérica), y 13 m de altitud en dirección ascendente (Hopkins 1918 en Daubenmire 1979). En las regiones templadas la actividad vegetal frecuentemente se detiene a principios del verano, mientras que la floración de plantas perennes termina a finales del verano; si el inicio del invierno se retrasa estas plantas tendrán un periodo breve de actividad vegetal y sin floración en el otoño (Grainger 1939 en Daubenmire 1979). Aunque el crecimiento de las raíces de algunas plantas perennes en las regiones templadas casi se interrumpe por el invierno, los brotes tienen periodos de descanso que normalmente pueden interrumpirse sin exponerse a bajas temperaturas (Daubenmire 1979). Diagramas climáticos Diagramas ombrotérmicos. A lo largo de este siglo se han propuesto diversos índices numéricos y gráficos para tratar de poner en evidencia las relaciones entre el clima y los seres vivos, en particular la vegetación. Los diagramas climáticos son muy útiles como expresiones gráficas del clima de cualquier territorio. Originalmente propuestos por Gaussen y popularizados por Walter y Leith, los diagramas ombroclimáticos pueden ser utilizados exitosamente para explicar los fenómenos bioclimatológicos (Rivas-Martínez 1987). El diagrama ombrotérmicos considera valores de precipitación, temperatura, balance hídrico y presencia de heladas. El promedio mensual de precipitación (P) y dos veces el de temperatura (2T), son graficados contra el tiempo (en meses), de tal manera que P=2T para cada punto en la escala de las ordenadas al origen. En el periodo seco, la línea de precipitación (P) se sitúa bajo la correspondiente a la temperatura (2T). Entre más grande sea el área entre las dos líneas, mayor será la aridez del clima (Bagnouls y Gaussen 1953 en Rivas-Martínez 1987). La figura 3 muestra la información clave para interpretar el diagrama ombrotérmico o climático con la siguiente información: nombre de la estación, altitud, número de años, temperatura media anual (T), precipitación media anual (P), heladas probables, periodo libres de heladas y periodo de actividad vegetal (PAV). 40 El bosque de coníferas GRAFICA 3. DIAGRMAS CLIMÁTICOS (1) (2) M (3) Años (6) T T: (4) P: (5) P (7) 300 200 100 (8) 80 40 60 30 (9) 40 20 20 10 0 0 (10) PAV 13 (11) H D E 12 F M A M J J A S O N D LEYENDAS: 1. Localidad de la estación 2. Altitud (m) 3. Años de los datos 4. Temperatura promedio anual (°C) 5. Precipitación promedio anual (mm) 7. Escala de precipitación 8. Línea de temperatura promedio mensual 9. Línea de precipitación promedio mensual 10. PAV: Período de actividad vegetal 12. H: Heladas 12. Período libre de heladas 13. Período de heladas probables. GRAFICA 4. DIAGRAMA CLIMÁTICO DE LA ESTACIÓN NO. 95 SIERRA SAN PEDRO MÁRTIR SAN PEDRO MÁRTIR, (BC) 2,080 m 7 Años T T: 10.7 °C P: 518.6 mm P 300 200 100 80 40 60 30 40 20 20 10 0 0 (10) PAV (11) H D E F M A M J J A S O N D Bioclimatología 41 A partir de datos disponibles para el periodo de 1984-1991, los cuales son insuficientes, se obtuvo el diagrama ombrotérmico de la estación No. 95 con el Bioclima (Alcaraz 1995), y que nos muestra un primer patrón climático de la Sierra de San Pedro Mártir. Así, el periodo de lluvias se inicia en el mes de octubre hasta el mes de abril (seis meses), mientras que en los meses de verano (julio-septiembre) se observan “picos” de lluvia de origen tropical. Con respecto a la temperatura, las bajas coinciden con el periodo invernal y las temperaturas altas con el verano. Aunque se presentan meses de heladas probables (diciembre a febrero), su ausencia durante el resto del año favorece la actividad vegetal (PAV) durante todo el año (Delgadillo 1995). Evapotranspiración y balance hídrico. Toda la humedad del suelo proviene de la precipitación, y sus características depende su efecto sobre la humedad edáfica (Deacon 1958 en Daubenmire 1979). En primer lugar, cuanto más lentos y moderados sean los aguaceros, mayor será el porcentaje de agua que se filtre en el suelo con relación a la que se pierde por escurrimiento. En segundo lugar, cuanto mayor sea la cantidad de agua que cae durante cualquier periodo lluvioso, mayor será la cantidad que se filtre más allá de la superficie de desecación. Por lo tanto, la existencia en climas secos de aguaceros que sumen varios centímetros de lluvia durante el verano puede que no produzca ningún efecto en el incremento del contenido de humedad del suelo, ya que cada lluvia es muy ligera y está demasiado distante de los otros eventos para provocar algún efecto acumulativo (Daubenmire 1979). La eficacia de la precipitación como fuente de humedad en el suelo se puede medir adecuadamente mediante estudios directos sobre el grado de penetración y de duración de la humedad en el suelo; estos métodos son muy complejos y lentos, aunque se logran resultados detallados. Entre tanto, se han realizado grandes esfuerzos para encontrar medios indirectos para evaluar este factor del medio ambiente de la planta (Dansereau 1957). El concepto de evapotranspiración es muy importante en ecología vegetal ya que evalúa perdidas de agua por evaporación del suelo y transpiración de las plantas, y contribuye a evaluar la efectividad de las lluvias. Los tipos de evapotranspiración son: a) potencial (si hubiera suficiente agua), lo que da una idea del estrés hídrico; y b) real, en función del potencial y del agua realmente disponible (Alcaraz 1999). 42 El bosque de coníferas Cada clima tiene una evapotranspiración potencial determinada principalmente por la energía recibida en forma de radiación solar, y en parte por el viento, ya que estos factores aumentan la temperatura y disminuyen la humedad relativa. Siempre que la humedad del suelo no sea muy limitante (lo que sucede con frecuencia), la evapotranspiración real iguala la evapotranspiración potencial. Ambas son similares en el caso de la vegetación densa tan distinta como los bosques o las praderas, a condición de que las copas de los árboles estén relativamente niveladas, el follaje tenga un reflejo similar y las actividades vegetativas de las plantas están cercanas al máximo (Dansereau 1957). El cálculo de la evapotranspiración se puede hacer con evapotransporímetros, y con el método de Blaney y Cridle (empírica) y el establecido por Thornthwaite (aproximación empírica) (Alcaraz 1999). Este último método se basa en las siguientes premisas: a) la temperatura está estrechamente relacionada con el abastecimiento de energía solar disponible para la evapotranspiración; b) la media de una temperatura mensual determinada está asociada a una mayor evapotranspiración cuando los días son más largos; c) el viento y la humedad atmosférica son de menor importancia, por lo que es posible hacer caso omiso de ellos sin peligro alguno para calcular la evapotranspiración, y d) el suelo tiene capacidad para almacenar aproximadamente 10 cm de lluvia. Por loque el significado biológico de la sequía no empieza de inmediato al iniciar la época cálida y sin lluvias (Dansereau 1957). Este cálculo permite evaluar el estrés hídrico a que se ven sometidas las plantas en los periodos más secos del año, que actuarán como limitantes, a los cuales deberán encontrarse adaptadas las diferentes especies que sobreviven en un clima dado (Álvarez 1992). Para el cálculo de la evapotranspiración de Thornthwaite, es usual aplicar un valor en la capacidad de almacenamiento de agua o reserva R, de 200 mm, que es bastante alto pero que, no obstante, no afecta a los resultados finales. Escoto (1994), estimó el balance hídrico de tres sitios de la Sierra (bosque bajo, bosque alto y pradera-Vallecitos) para el periodo 1989-1992, para lo cual consideró la humedad del suelo, el escurrimiento en pequeñas parcelas y el drenaje profundo en minilisímetros y calculó la evapotranspiración (ET) para la capa de 0-80 cm. La ET de invierno fue aproximadamente 40% de la precipitación (501 ± 17), y la de verano 125% (191 ± 100). El suelo de Bioclimatología 43 los bosques es de textura gruesa y la roca se encuentra a una profundidad entre 60 y 80 cm, y por lo tanto tienen baja capacidad de almacenamiento de agua entren 140 y 980 mm, sin embargo el suelo de la pradera es profundo y de textura mediana, la capacidad de almacenamiento de agua es en la capa de 0-150 cm, y alcanza un valor de almacenamiento de 230 mm. La gráfica 5 muestra un diagrama de Thornthwaite con el balance hídrico de la estación metereológica No. 95 a partir de los datos disponibles. Con líneas verticales se indica el periodo de precipitación y acumulación de la reserva, las líneas punteadas se refieren a la disponibilidad de agua como reserva para las plantas, y en blanco aparecen los meses con déficit hídrico. En este caso, los datos de la estación No. 95 y que corresponden al sitio de bosque bajo de Escoto (1994), muestran una precipitación de 341.6 mm en el periodo invierno-primavera (diciembre-mayo) y una ETR de 179 mm, mientras que la precipitación para periodo verano-otoño fue de 183.3 mm y una ETR de 287.8 mm (cuadro 5). FIGURA 5. BALANCE HÍDRICO DE LA ESTACIÓN METEREOLÓGICA NO. 95, SIERRA SAN PEDRO MÁRTIR 44 El bosque de coníferas Bioclimatología 45 5.4 F E 4.1 64.0 19.3 19.3 44.7 170.1 0.0 0.0 99.5 14.6 14.6 84.9 125.4 0.0 0.0 F 5.9 M 106.6 25.2 25.2 0.0 200.0 0.0 0.0 M 8.3 A 8.2 39.0 39.0 -30.8 169.2 0.0 0.0 A 10.6 M 6.1 56.3 56.3 -50.2 119.0 0.0 0.0 M 16.3 J 0.7 91.1 91.1 -90.4 28.6 0.0 0.0 J Altitud 2898 m; P: 518.6 mm: T: 18.7° C 18.2 J 57.5 105.9 86.1 -28.6 0.0 19.8 0.0 J 16.9 A 37.9 92.4 37.9 0.0 0.0 54.5 0.0 A 15.2 S 30.0 74.0 30.0 0.0 0.0 44.0 0.0 S 11.1 O 7.0 49.3 7.0 0.0 0.0 42.3 0.0 O 9.2 N 43.9 35.7 35.7 8.2 8.2 0.0 0.0 N 6.7 D 57.2 24.9 24.9 32.3 40.5 0.0 0.0 D Superávit P. Precipitación ETP. Evaporación potencial ETR. Evaporación real VR. Incremento de agua en la reserva R. Reserva DEF. Déficit SUP. T P ETP ETR VR R DEF SUP E San Pedro Martir (BC) CUADRO 5. DATOS DE LA EVAPOTRANSPIRACIÓN (ET) DE LA ESTACIÓN METEREOLÓGICA NO. 56, SIERRA DE SAN PEDRO MÁRTIR, BAJA CALIFORNIA Pisos bioclimáticos Se entiende por pisos bioclimáticos cada uno de los tipos o espacios termoclimáticos que tienen lugar en una cliserie altitudinal o latitudinal (Rivas-Martínez 1987), esto es, la serie de asociaciones vegetales determinadas por la sucesión de climas en sentido altitudinal o latitudinal (FontQuer 1979). En la práctica, los pisos bioclimáticos se reconocen mediante las comunidades vegetales que presentan correlaciones con determinados intervalos climáticos. La altitud y latitud son factores que influyen fuertemente en la distribución de las especies, debido a que inducen a una verdadera zonación (Dansereau 1957). Dentro de un área homogénea, y desde el punto de vista del macrobioclima, los accidentes del relieve y la topografía confieren una originalidad a muchos territorios geográficos. La distribución de la vegetación en pisos, bandas o cinturones en función del cambio de la temperatura con la altitud, se conoce como zonación altitudinal. Este hecho es especialmente claro en el caso de las montañas, donde el descenso de la temperatura media anual es de unos 0.65 °C por cada 100 m de ascenso en altitud. Este tipo de cambios se da también en sentido latitudinal, aunque en este caso son más suaves, ya que se necesita aproximadamente de unos 100 km hacia el polo geográfico más próximo para que se de un descenso similar en la temperatura media anual. Junto a esta zonación altitudinallongitudinal de la vegetación, la variación de las precipitaciones y/o de su efectividad supone un segundo factor que induce también cambios en el manto vegetal (Rivas-Martínez 1987). La combinación de pisos bioclimáticos se refleja en la distribución tanto de plantas como de comunidades vegetales, las cuales en definitiva se manifiestan paisajísticamente como pisos de vegetación, en grandes territorios, como formaciones fisionómicas. Numerosas plantas y comunidades están perfectamente ceñidas a una determinada combinación termoombroclimática y, en consecuencia, pueden usarse como bioindicadores de pisos, ombroclimas o formaciones y, a su vez, de unidades biogeográficas (Delgadillo 1999). Cada región biogeográfica tiene pisos bioclimáticos especiales y un componente térmico (termotipo) y otro ómbrico (ombrotipo). El termotipo se puede reconocer mediante el del índice de termicidad compensado o, 46 El bosque de coníferas con bajas temperaturas, del índice de temperaturas positivas, mientras que el ombrotipo se puede determinar a partir del índice ómbrico (árido, semiárido, seco, subhúmedo húmedo, hiperhúmedo) (Alcaraz 1999). Los valores térmicos se calculan mediante el índice de termicidad: It = (T+m+M) 10, donde «T» es la temperatura media anual, «m» la temperatura media de las mínimas del mes más frío, y «M» la temperatura media de las máximas del mes más frío. La bondad de este índice se basa en que el valor de las mínimas del mes más frió (m) y su corrección de acuerdo con su duración a lo largo del día con la media de las máximas también del mes más frío del año (M), opera como factor limitante en la ley del mínimo. La importancia y peso que confiere esta fórmula o índice de termicidad al frío estacional o altitudinal ha resultado ser de gran utilidad y elevada correlación con la distribución de plantas y sus fitocenosis en la tierra (RivasMartínez 1987). Los pisos bioclimáticos colino, montano, subalpino y alpino son caracteristicas de la región Pacífico-Norteamericana, mientras que para la región Californiana se han reconocido los pisos inframediterráeo, termomediterráneo, mesomediterráneo, supramediterráneo, oromediterráneo y crioromediterráneo. Para la región Xerofítico-Mexicana son característicos los pisos termotropical, mesotropical y supratropical (Peinado et al. 1994b). En la zona de clima mediterránea de California y noroeste de Baja California existe un gradiente térmico latitudinal y altitudinal que permite identificar los siguientes pisos bioclimáticos: inframediterráneo, termomediterráneo, mesomediterráneo y supramediterráneo, mientras que el mesotropical se observa en la zona de clima tropical-desértico situada al este de la sierra. Para cada piso se han distinguido varios ombroclimas, los cuales se delimitan por intervalos de la precipitación anual en mm (Peinado et al. 1994b) (figura 6). Los anteriores pisos bioclimáticos termotropical, mesotropical e inframediterráneo presentan los siguientes ombroclimas: semiárido (P=200400), seco (P=400-650) y árido (P=50-200) respectivamente, mientras que en los pisos termomediterráneo, mesomediterráneo y supramediterráneo existen los ombroclimas semiárido (P=160-350), seco (P= 350-500) y subhúmedo (P= 500-900) (Peinado et al. 1994b, Delgadillo 1995). El carácter más distintivo de la Sierra de San Pedro Mártir es la asimetría climática existente entre sus vertientes occidental y oriental. En la primera, Bioclimatología 47 son claras las influencias del océano Pacífico que conduce a la aparición de un clima claramente mediterráneo de veranos secos y calurosos e inviernos relativamente húmedos y fríos. Mientras que la vertiente oriental de la Sierra, expuesta al clima árido del Golfo de California, queda en la sombra de lluvias y muestra diagramas ombrotérmicos típicamente tropicaldesérticos (Peinado et al. 1994b). En un transecto que va desde el Océano Pacífico hacia el Golfo de California, pasando por la Sierra de San Pedro Mártir, se han identificado cinco de los seis pisos bioclimáticos presentes en la península de Baja California. En su cara oeste se encuentran los pisos inframediterráneo, termomediterráneo, mesomediterráneo, supramediterráneo; mientras que en las laderas orientales encontramos además del mesomediterráneo el piso mesotropical. Sin embargo, los bosques de coníferas en la sierra sólo se presentan los pisos mesomediterráneo y supramediterráneo (Delgadillo 1995) (grafica 7). FIGURA 6. PISOS DE VEGETACIÓN DE BAJA CALIFORNIA Inframediterráneo Termomediterráneo Mesomediterráneo Supramediterráneo Mesotropical Termotropical Fuente: Peinado et al. 1994b, Delgadillo 1995. 48 El bosque de coníferas FIGURA 7. DISTRIBUCIÓN APROXIMADA DE LOS PISOS BIOCLIMÁTICOS Y VEGETACIÓN 3,000 m Supramediterráneo Supramediterráneo 2,000 m Mesomediterráneo Mesomediterráneo 3 1,500 m Termomediterráneo Mesotropical L. Monje ‘95 3 500 m Inframediterráneo 3 Mar de Cortés 2 SECTOR MARTIRENSE Inframediterráneo = Termomediterráneo = Mesomediterráneos = Suprarmediterráneo = Mesotropical = SECTOR SANFELIPENSE Matorrales costeros suculentos (1, 2) Chaparrales y encinares (3) Chaparrales y bosques de pino piñonero Bosques de Pinus jeffreyi con comunidades de Abies concolor y Calocedrus decurrens (7) Matorral micrófilo (desierto de San Felipe) Fuente: Delgadillo 1995. Piso mesomediterráneo El piso mediterranáneo está dominado por un chaparral de montaña (Delgadillo 1998) y coníferas heliófilas de comunidades de pinos piñoneros (Pinus quadrifolia y P. monophylla), que forman bosquetes abiertos entre los 1,300 y 2,000 m. La escasez de precipitaciones en el noroeste de Baja California impide por lo general la existencia de bosques de esclerófilos en este piso, desarrollándose en su lugar bosques xerofíticos abiertos dominados por P. quadrifolia (pino piñonero) y Adenostoma sparsifolium (chamizo vara roja). Los ombroclimas oscilan entre el árido (únicamente en la vertiente oriental) y el semiárido (foto 7). Un factor importante en la distribución del chaparral es el efecto orográfico del aire frío adiabático con el incremento de la elevación, que Bioclimatología 49 FOTO 7. PISO BIOCLIMÁTICO MESOMEDITERRÁNEO EN LA VERTIENTE OESTE, CON VEGETACIÓN DE CHAPARRAL produce un descanso de la temperatura de crece con el incremento de la elevación. Otro factor de importancia local en el sur de California lo representa la influencia de los llamados «vientos de Santa Ana», los cuales resultan de una alta presión en el interior de los Estados Unidos de América, llevando aire seco desértico hasta la costa. Este tipo de vientos puede exceder los 100 km/hr y traer altas temperaturas y baja humedad, siendo comunes en primavera y otoño. Algunos de los fuegos más devastadores ocurren bajos estas condiciones climáticas (Keeley y Keeley 1989). El sotobosque de P. quadrifolia presenta comunidades de Adenostoma sparsifolium y Juniperus californica (véase asociación fitosociológica Adenostomo sparsifolii-Pinetum quadrifoliae). Al parecer esta comunidad, por acción del fuego, ha evolucionado hacia una facies de A. sparsifolium en la que los pinos piñoneros quedan como individuos aislados o llegan a faltar por completo. En el límite altitudinal de estos pinares se mezclan elementos supramediterráneos como Pinus jeffreyi y Arctostaphylos patula (manzanita). Minnich (1987) refiere que individuos aislados de P. quadrifolia 50 El bosque de coníferas alcanzan los 2,700 m cerca del Observatorio Nacional. Peinado et al. (1994) han señalado que este pino es un árbol muy escaso en el piso supramediterráneo; Delgadillo (1999) no lo reporta para esta zona. Los bosques abiertos de Pinus monophylla y de algunos Juniperus son un tipo de vegetación común en todas las fachadas orientales hacia los desiertos, desde San Pedro Mártir (Juniperus californica) hasta las Montañas Rocosas (J. osteosperma), bordeando altitudinalmente formaciones tanto del desierto neotropical (Desierto de Sonora) como holártico (Gran Cuenca) (Peinado et al. 1994b). En la Sierra de San Pedro Mártir estos bosques se presentan en su vertiente oriental bajo condiciones ombrotérmicas árido-semiárido, formando un cinturón de vegetación entre los matorrales mesotropicales de Agave deserti y los bosques densos supramediterráneos de Pinus jeffreyi y Abies concolor, que los sustituyen altitudinalmente (Peinado et al. 1994b). La distribución de P. monophylla y P. quadrifolia en Baja California refleja los requerimientos ombrohídricos de ambos taxa. Aunque los dos son mesomediterráneos, el primero habita preferente bajo ombroclima áridosemiárido, de ahí que las mejores poblaciones ocupen de forma más o menos continua los flancos orientales de las sierras de Juárez y San Pedro Mártir. Por su parte, P. quadrifolia habita preferentemente bajo ombroclima semiárido o seco inferior, por lo que los mejores bosques se encuentran en el piso mesomediterráneo, pero en exposiciones oeste donde reciben la humedad del Pacífico. Ambas especies pueden estar en contacto, sobre todo en los límites altitudinales del escarpe oriental y norte de San Pedro Mártir (Cerro Venado Blanco, Rancho El Picacho), por debajo de los bosques supramediterráneos de coníferas, donde se inicia el efecto de sombra de lluvia y se produce la transición entre los ombroclimas seco y semiárido. En la mayor parte del piso mesomediterráneo del suroeste de Norteamérica la vegetación climácica está constituida por encinares de Quercus agrifolia, los cuales necesitan un ombroclima al menos seco. Estos encinares tienen su límite en el piso supramediterráneo por encima de los 1,800 m del lado oeste de San Pedro Mártir, apareciendo en zonas riparias o galerías en fluvisoles con ligera hidromorfía. Bioclimatología 51 Piso supramediterráneo En términos bioclimáticos el ombroclima donde se encuentran los bosques de la clase Arctostaphylo-Pinetea lambertianae oscila entre el seco y el húmedo para las zonas de más baja precipitación ocupadas por los bosques abiertos. El rango de precipitaciones dentro del cual se desarrollan estos bosques es muy variable. Barbour (1988), Major y Taylor (1988) y Hastings y Turner (1965), han reportado valores de precipitación de estaciones situadas en el área de los bosques de la clase que oscilan entre los 500 mm en la Sierra de San Pedro Mártir, hasta más de 1,200 en Sierra Nevada y el sur de las Cascadas en California. A estas medidas pluviométricas hay que añadirles el aporte extraordinario de las criptoprecipitaciones y del agua procedente de la nieve en el deshielo, en buena parte causante del mantenimiento de la eleva biomasa de estos bosques incluso en sus áreas más secas (Pase 1982). Este piso de vegetación se encuentra en la Sierra de San Pedro Mártir por arriba de los 2,000 msnm, donde presenta grandes formaciones forestales dominadas por árboles de gran talla de Pinus jeffreyi, P. lambertiana, P. contorta var. murrayana, Abies concolor y Calocedrus decurrens, que constituyen la vegetación climácica de este piso hasta Sierra Nevada, California. A diferencia de los bosques de Sierra Nevada, en la Sierra de San Pedro Mártir falta un encino caducifolio, Quercus kelloggii, lo que probablemente se deba a la escasa precipitación. Por el contrario, Quercus chrysolepis es un encino esclerófilo que en Sierra Nevada se encuentra preferentemente en el piso mesomediterráneo, mientras que en la Sierra de San Pedro Mártir es una especie exclusiva del piso supramediterráneo (Peinado et al. 1994b). El elemento dominante en el piso supramediterráneo es P. jeffreyi (foto 8), que, ocasionalmente, puede introducirse en piso mesomediterráneo en zonas riparias o galerías dominadas por Q. agrifolia. Por otra parte, P. lambertiana es menos frecuente, aunque se vuelve progresivamente dominante a mayores altitudes y en exposiciones al norte. Abies concolor es común por encima de los 2,200 m y, al igual que P. lambertiana, prefiere exposiciones al norte, con mayor humedad y umbría, incluyendo las partes altas del Picacho del Diablo. Pinus contorta var. murrayana es un taxa fundamentalmente oromediterráneo en Sierra Nevada, sin embargo en la Sierra de San Pedro Mártir se presenta en los márgenes de las praderas (por ejem- 52 El bosque de coníferas plo, Vallecitos), donde hay una gran compensación de humedad. En cuanto a Calocedrus decurrens, este tipo de cupresáceas representa el árbol más raro (algunos de hasta 45 m de altura) y prefiere suelos húmedos (camino a La Tasajera). Otros árboles comunes sobre arroyos temporales o de agua permanente son Populus tremuloides y Salix spp. (Delgadillo 1999). FOTO 8. PISO BIOCLIMÁTICO SUPRAMEDITERRÁNEO CON BOSQUE DE CONÍFERAS Bioclimatología 53 54 El bosque de coníferas Capítulo 4. Fitogeografía Los grandes rangos o jerarquías que se aceptan en fitogeografía son reino, región, provincia y sector. Existe un práctico acuerdo en considerar a todas estas unidades como territorios de superficie continua (excepción hecha de las islas), que incluyan los accidentes orográficos que puedan existir en su área (Rivas-Martínez 1987). Salvo raras excepciones, en su mayoría provocadas por cambios en el sustrato, las fronteras de las unidades no son abruptas, de forma que unidades vecinas comparten ecotonos más o menos extensos. Por lo general, son los macroclimas los que limitan las unidades de mayor rango (reinos, regiones), mientras que son factores topográficos, geológicos, edáficos o climáticos regionales los que delimitan provincias y sectores. En cualquier caso, es la distribución de las plantas y comunidades la que marca la extensión y los límites de las unidades fitogeográficas (Peinado et al. 1994a, Delgadillo 1995). La unidad fitogeográfica de más alto rango es el reino y está caracterizado por la existencia de zonobiomas, de grandes formaciones vegetales, de unidades fitosociológicas de nivel más alto (clases) y de táxones al nivel de familias u ordenes. En Baja California se reconocen dos reinos: Holártico y Neotropical, cuyas diferencias florísticas son bien conocidas (Peinado et al. 1994a). Las regiones se dividen en provincias y para su discriminación se han utilizado varios criterios. La provincia posee sólo algunos de los pisos bioclimáticos que caracterizan a su correspondiente región, aunque en esta última la cliserie altitudinal se presenta completa. Por ejemplo, la cliserie altitudinal de la región californiana va desde el piso inframediterráneo al crioromediterráneo, pero esta cliserie no se presenta íntegramente en ninFitogeografía 55 guna de sus provincias. Así, en la provincia Martirense están representados los pisos infra, termo, meso y supramediterráneo, siendo el primero de ellos una característica de esta provincia por lo que, evidentemente, los táxones propios de este piso constituyen por añadidura un elemento florístico de peso para independizarla fitogeográficamente. Al igual que los pisos, el factor que más condiciona la distribución de las comunidades seriales es el ombroclima; por ello, los ombroclimas pueden ser esenciales a la hora de diferenciar provincias que contengan la misma cliserie altitudinal. A nivel fitosociológico las provincias están caracterizadas por asociaciones climácicas, alianzas e incluso provincias, especialmente aquellas ricas en flora por su aislamiento biogeográfico. Por lo demás, las provincias poseen algunos géneros endémicos y un buen conjunto de especies endémicas, entre las que, por lo general, se cuentan algunos paleoendemismos. Dentro de una misma provincia el sector puede caracterizarse de diferentes formas y, desde el punto de vista fisiográfico, los sectores suelen coincidir con grandes unidades geomorfoestructurales tales como grandes sierras o extensas superficies zonobioclimáticas o edafológicamente importantes. Peinado et al. (1994a) reconocen en la península de Baja California las siguientes unidades fitogeográficas (figura 8). Región californiana Esta región ocupa la mayor parte del estado de California y una pequeña porción del sur de Oregón y se extiende hacia el sur hasta el noroeste de Baja California, incluyendo la Sierra de San Pedro Mártir. Esta región alcanza el paralelo 30°, donde inicia la región xerofítica-mexicana (provincia Bajocaliforniana). Las vertientes orientales de las Sierras Juárez y de San Pedro Mártir no pertenecen en su mayor parte a esta región, sino a la Xerofítico-Mexicana, principalmente por debajo de los 1,000 m (Peinado et al. 1994a). La Sierra de San Pedro Mártir se encuentra dentro de la región Californiana, la cual está dividida a su vez en las siguientes provincias: a) Nevadense; b) Californiano-Septentrional; c) Californiana-Meridional, y d) Martirense. En este texto sólo se hará la descripción de ésta última provincia ya que la Sierra de San Pedro Mártir es parte de ella. 56 El bosque de coníferas FIGURA 8. FITOGEOGRAFÍA DE BAJA CALIFORNIA Reino Holartico Región Californiana I. Provincia californianomeridional (Sector Diegano) II.- Provincia martirense IIa. Sector Juarezense IIb. Sector Martirense Reino Neotropical Región Xerofítico-mexicana III. Provincia bajocaliforniana IIIa. Sector vizcaíno IIIb. Sector Angelino-Loretano IIIc. Sector Magdalenense IV. Provincia sanlucana (Sector Sanlucaño) V. Provincia colorada (Sector Sanfelipense) Fuente: Peinado et al. 1994a. Provincia Martirense En el noroeste de la península de Baja California la región Californiana ocupa alrededor de 24,500 km.². La provincia Martirense abarca la mayor parte de este territorio donde existen, al menos, dos grandes unidades morfoestructurales, las Sierras de San Pedro Mártir y Juárez. Éstas están perfectamente caracterizadas desde el punto de vista florístico, ya que albergan un importante conjunto de endemismos a nivel específico y subespecífico, además de presentar una cliserie altitudinal diferente. Ambas sierras, junto a las sierras costeras del Pacífico, se consideran como dos sectores independientes (Peinado et al. 1994a). Los límites septentrionales de esta provincia se establecen desde el sur de la Bahía de San Diego hasta el sur de la Mesa de Otay, dirigiéndose hacia el oeste por la cuenca del río de Las Palmas y el borde meridional de la falla Fitogeografía 57 La Rumorosa-Las Palmas, hasta alcanzar su límite nororiental en las cumbres de La Rumorosa. Por el este, la provincia Martirense limita con la región Xerofítico-Mexicana, donde la frontera biogeográfica entre ambas son las vertientes orientales de las Sierras Juárez y de San Pedro Mártir por debajo de los 1,300-1,100 m aproximadamente, cota inferior alcanzada por los bosques semiáridos mediterráneos de Pinus monophylla y Juniperus californica. El límite meridional de la provincia se establece alrededor del paralelo 30 °N, unos kilómetros al sur del arroyo El Rosario, donde inicia el sector Vizcaíno de la provincia Bajocaliforniana (Peinado et al. 1994a). Aproximadamente un centenar de especies de plantas son endémicas de esta provincia y una buena parte son esquizoendemismos orófilos supramediterráneos. Estos endemismos probablemente se han originado como consecuencia del aislamiento geográfico de las cumbres de la Sierra de San Pedro Mártir con respecto a otras unidades de la región Californiana en las que existe el piso supramediterráneo. Una buena parte de los táxones Californiano-Meridionales que no alcanzan la provincia Martirense son claramente plantas que requieren mayor humedad; esto explica la falta de elementos que resultan tan ubicacuos y conspicuos en las vecinas montañas peninsulares y transversales como Umbellularia californica, Quercus kelloggii y Pinus ponderosa (Peinado et al. 1994a). En la provincia se distinguen dos sectores: Juarezense y Martirense; este último se extiende por la mitad sur de la provincia, inicia cerca de Punta Colonet y se dirige hacia el este hasta el Paso de San Matías, cuya falla constituye la frontera biogeográfica con el sector Juarezense. La escasa altura de la Sierra de Juárez impide las precipitaciones orográficas que permiten la existencia de bosques supramediterráneos de Abies concolor, Pinus lambertiana, Pinus contorta o Calocedrus decurrens. 58 El bosque de coníferas Capítulo 5. Composición florística Flora El bosque de coníferas en nuestra región tiene elementos florísticos que merecen su consideración a cualquier nivel. Howell (1957 en Axelrod 1978) refiere que el noroeste de la península es una de las regiones con respecto a su riqueza florística. La flora de la península de Baja California cuenta con aproximadamente 3,100 taxa (especies, subespecies y variedades), incluidos en 155 familias botánicas. De manera particular, las plantas de la Sierra de San Pedro Mártir han sido recolectadas por diferentes botánicos, quienes han contribuido a conocer el componente florístico del bosque de coníferas (Nelson y Goldman 1905-1906 en Goldman 1916,Wiggins 1980, Moran y Thorne 19671986, Delgadillo 1981-2002, Rebman 1994-2002). Passini et al. (1989) recolectaron en el interior del bosque y reportaron 154 táxones, repartidos en 85 géneros y 44 familias. Por otra parte, Minnich y Franco-Vizcaíno (1997) refieren la presencia de 116 especies de plantas con flor en la pradera de Vallecitos, pero no hacen referencia a su colecta ni presentan un listado florístico. Moran y Thorne (1967-1986) recolectaron en la mayor parte de la zona boscosa de la Sierra y registrando 58 familias, 175 géneros y 331 taxa, de los cuales 29 fueron variedades y 23 subespecies. Delgadillo (1999) identifico para la Sierra de San Pedro Mártir, incluyendo la vegetación de chaparral de contacto, un total de 74 familias, 286 géneros y 735 taxa: 584 son especies y 251 con categorías infraespecíficas (subespecie y variedad). Las familias más numerosas fueron: Asteraceae, Composición florística 59 Poaceae, Fabaceae, Scrophulariaceae, Polygonaceae, Brassicaceae, Cyperaceae, Polemoniaceae y Lamiaceae (cuadro 6). A partir de los registros de diferentes autores que han colectado en la Sierra de San Pedro Mártir, así como la base de datos del Herbario BCMEX, de la Facultad de Ciencias (UABC), se estima una flora exclusiva al bosque de coníferas de aproximadamente 580 taxa, incluidos 151 subespecies y variedades, en 72 familias (ver anexo II, catálogo florístico). CUADRO 6. LAS DIEZ FAMILIAS CON EL MAYOR NÚMERO DE TAXA EN LA SIERRA SAN PEDRO MÁRTIR, BAJA CALIFORNIA, MÉXICO FAMILIAS Asteracea Poaceae Fabaceae Scrophulariaceae Cyperaceae Rosaceae Polygonaceae Brassicaceae Lamiaceae Polemonaceae Total TAXA % 83 66 38 26 23 22 21 21 15 14 329 14.3 11.3 6.5 4.4 3.9 3.7 3.6 3.6 2.5 2.4 56.7 Formas biológicas Cruz (2001) señala que en la Sierra de San Pedro Mártir la forma biológica más abundante fueron las herbáceas con un 75% de las especies, siguiéndole con una notable diferencia las arbustiva con 19%. El porcentaje restante está dividido entre ellos árboles, suculentas y sufrutescentes. Las familias con mayor número de herbáceas reportadas son: Asteraceae, Poaceae, Fabaceae, Polygonaceae y Brassicaceae; los géneros con el mayor número de taxa y su forma biológica se muestra en el cuadro 7 (Delgadillo 1999). Mientras que las familias con un mayor índice de arbustos y 60 El bosque de coníferas sufrutescentes son: Scrophulariaceae, Rosaceae, Polygoniaceae, Rhamnaceae y Ericaceae. CUADRO 7. GÉNEROS CON EL MAYOR NÚMERO DE TAXA Y SU FORMA BIOLÓGICA EN LA SIERRA DE SAN PEDRO MÁRTIR, BAJA CALIFORNIA, MÉXICO GÉNEROS Juncus (Cyperaceae) Muhlenbergia (Poaceae) Eriogonum (Polygonaceae) Lotus (Fabaceae) Erigeron (Asteraceae) Phacelia (Hydrophyllaceae) Galium (Rubiaceae) Haplopappus (Asteraceae) Lupinus (Fabaceae) Mimulus (Scrophulariaceae) Quercus (Fagaceae) Eleocharis (Cyperaceae) Camissonia (Onagraceae) Potentilla (Rosaceae) Ceanothus (Rhamnaceae) Pinus (Pinaceae) Cirsium (Asteraceae) Astragalus (Fabaceae) Arctostaphylos (Ericaceae) Rhamnus (Rhamnaceae) Descurainea (Brassicaceae) TÁXONES 14 14 12 11 9 9 8 8 7 7 7 7 6 6 6 6 6 5 5 5 5 FORMA BIOLÓGICA Herbácea Herbácea Sufrutescente Herbácea Herbácea Herbácea Herbácea Sufrutescente Herbácea Herbácea Arbustiva Herbácea Herbácea Herbácea Arbustiva Árbol Herbácea Herbácea Arbustiva Arbustiva Herbácea Los árboles están restringidos a las familias Pinaceae, Cupressaceae, Fagaceae y Salicaceae. El cuadro 8 contiene los porcentajes de las formas biológicas, mientras que el 9 reune los taxa predominantes. ENDEMISMO La importancia y singularidad de la flora mexicana recaen en el número total de especies, pero sobre todo, en su riqueza de endemismos y su alto Composición florística 61 CUADRO 8. PORCENTAJE DE LAS FORMAS BIOLÓGICAS DE LA FLORA VASCULAR SIERRA DE SAN PEDRO MÁRTIR FORMAS EN LA % BIOLÓGICAS Hierbas perennes Hierbas anuales Arbustos Árboles Suculentas Sufrutescentes Parásitas Acuáticas Saprofitas Subarbustos Trepadoras 39.2 31.1 18.2 2.8 2.7 1.9 1.0 1.0 0.7 0.6 0.4 CUADRO 9. ALGUNOS DE LOS TAXA PREDOMINANTES EN EL BOSQUE DE CONÍFERAS EN LA SIERRA DE SAN PEDRO MÁRTIR Arbóreas Sufrutescentes Abies concolor Calocedrus decurrens Cupressus montana Pinus contorta var. murrayana Pinus coulteri Pinus jeffreyi Pinus lambertiana Pinus monophylla Pinus quadrifolia Populus tremuloides Quercus chrysolepis Quercus peninsularis Quercus turbinella Salix lasiolepis Eriogonum wrigthii Ericameria arborescens ssp. peninsularis Haplopappus martirensis Salvia pachyphylla Herbáceas Astragalus gruinus Castilleja martinii Euphorbia palmeri Linanthus melingii Monardella linoides Phlox austromontana Potentilla spp. Monardella macrantha Thalictrum fendleri (Continúa) 62 El bosque de coníferas CUADRO 9. ALGUNOS DE LOS TAXA PREDOMINANTES EN EL BOSQUE DE CONÍFERAS EN LA SIERRA DE SAN PEDRO MÁRTIR Arbustivas Amorpha apiculata Amorpha californica Arctostaphylos patula ssp. platyphylla Arctostaphylos peninsularis Arctostaphylos pringlei ssp. drupacea Berberis higginsae Ceanothus cordulatus Ceanothus leucodermis Ceanothus spinosus Garrya grisea Holodiscus microphyllus Philadelphus microphyllus Rhamnus californica ssp. tomentella Symphoricarpus parishii Arceuthobium campypodum Arceuthobium Cuscuta californica Cuscuta ceanothi Phoradendron densum Phoradendron lobrocedi Phoradendron macrophyllum Phoradendron pauciflorum Phoradendron villosum ssp. villosum Saprofitas Sarcodes sanguinea Pterospora Andrómeda Parásitas Arceuthobium americanum Arceuthobium californicum porcentaje lo cual se explica por la antigüedad de la flora mexicana, así como por su grado de aislamiento ecológico. Aunque existe una cierta relación florística entre las zonas áridas mexicanas y las estadounidenses, las primeras presentan una diferencia climática importante que las mantiene “ecológicamente separadas” (Rzedowski 1992). Wiggins (1980) menciona que 23.2% del total de la flora de Baja California es endémica, sin presencia de familias endémicas. El número de especies endémicas para el bosque de coníferas de las Sierras de Juárez y de San Pedro Mártir es notorio debido tal vez a su relativo aislamiento y condiciones ambientales específicas. Indiscutiblemente, estas dos áreas boscosas tienen todavía muchos táxones que pueden ser nuevos para la ciencia y/o registros biogeográficos (Delgadillo1998). Oberbauer (1996) refiere 12 y 19 taxa endémicos para las Sierras de Juárez y de San Pedro Mártir, respectiva- Composición florística 63 mente. Por su parte Minnich et al. (1997), consideran 18 especies endémicas al bosque de San Pedro Mártir, sin embargo se reducen a 16 ya que la distribución de dos de ellas se extiende hacia la Sierra de Juárez y sur de California, respectivamente. Delgadillo (1999) refiere para el bosque de coníferas y sus límites con el chaparral de la Sierra de San Pedro Mártir 25 taxa endémicos, que representan 4.3% del total de 580 taxa registrados. Sobresale Cupressus montana como la única especie arbórea y restringida en cañadas; las restantes formas biológicas son: herbáceas (15), sufrutescentes (6), arbustos (2) y suculentas (1) ( (cuadro 10). Consideraciones florísticas especiales Pinus ponderosa. Debido al aspecto similar de Pinus jeffreyi y Pinus ponderosa, mucho se ha discutido la posible ocurrencia de P. ponderosa en Baja California, ya que existen quienes la citan en sus trabajos (Martínez 1945 en Farjon et al. 1997, Inventario Forestal de Baja California 1968,Wiggins 1980 y Eguiluz 1985), como aquellos que no han reconocido individuos que reúnan características de esta especie en los bosques de pinos de ambas sierras (Goldman 1916; Moran 1977, Brown 1982, Thorne y Moran 1986, Thorne 1985, 1986, 1987, Beaucahmp 1986, y Minnich 1987, Peinado et al. 1994b, Delgadillo 1998). La cita de P. ponderosa Baja California por Wiggins (1980) no ha sido comprobada por ningún otro botánico, ni existen testimonios de herbario. En el Inventario Forestal del Estado de Baja California de 1968 (SARH 1977) se reporta a Pinus ponderosa para las sierras de Juárez y de San Pedro Mártir, sin embargo los autores de este inventario seguramente se confundieron al identificar como P. ponderosa a los pinos de “corteza rojiza” y P. jeffreyi aquellos con “corteza negra”, característica insuficiente para la identificación de un taxa. Lo anterior indica la edad del árbol: corteza negra en los individuos jóvenes y corteza roja en los viejos. Una manera fácil de identificar a P. jeffreyi es su olor a piña-vainilla en su corteza (Delgadillo1998). Goldman (1916) fue el primero en discutir la ocurrencia de P. ponderosa en la Sierra de San Pedro Mártir, al examinar características fácilmente separables de conos pequeños, grandes e intermedios de árboles próximos, sin embargo abandonó este carácter como falto de confianza, agregando que un estudio de estos pinos puede demostrar la ocurrencia de dos formas, tal y como sucede en el sur de California 64 El bosque de coníferas CUDRO 10. RELACIÓN DE LOS TAXA ENDÉMICOS REGISTRADOS EN LA SIERRA DE SAN PEDRO MÁRTIR, BAJA CALIFORNIA, MÉXICO TAXA Allium eurotophyllum Amorpha apiculata Astragalus gruinus Brickellia sessile Cordylanthus involutus Cupressus montana Draba corrugata var. demareei Dudleya pauciflora Ericameria parishii var. peninsularis Ericameria martirensis Eriogonum elongatum Galium diabloense Galium martirense Galium wigginsii Garrya grisea Haplopappus pulvinatus Hedeoma martirense Hemizonia martirensis Heterotheca martirensis Lesquerella peninsularis Ophiocephalus angustifolius Poliomintha conjunctrix Stephaeromeria martirensis Stephanomeria monocephala Trifolium wigginsii FORMA BIOLÓGICA Herbácea Arbustiva Herbácea Herbácea Herbácea Árbol Herbácea Suculenta Sufrutescente Sufrutescente Herbácea Sufrutescente Herbácea Herbácea Arbustiva Sufrutescente Herbácea Sufrutescente Herbácea Herbácea Herbácea Sufrutescente Herbácea Herbácea Herbácea Duffield y Cumming (1974), al preguntarse también la posible presencia de P. ponderosa en Baja California, mencionan que de acuerdo a su experiencia los árboles con tamaños intermedios del cono, la mayoría de ellos observados con mejor ventaja en árboles vivos, son casi sin excepción fácilmente reconocidos como pertenecientes a cualquiera de las dos especies, P. ponderosa o P. jeffreyi, pero este reconocimiento depende de otras características y no sólo del tamaño del cono. La distinción entre P. ponderosa y P. jeffreyi debe continuar abierta ante más evi- Composición florística 65 dencias acumuladas y analizadas sobre distribución, genética y estudios taxonómicos. Análisis químicos de las oleresinas del xilema de P. jeffreyi revelan que las turpentinas consisten casi completamente (90-95%) de heptanos, mientras que en P. ponderosa generalmente están ausentes en sus compuestos. La presencia de las turpentinas ayuda a una identificación rápida en campo ya que en días cálidos los troncos presentan un olor dulce a miel o piña-vainilla (Perry 1991). Pseudotsuga macrocarpa. También se ha cuestionado la presencia de esta especie característica de los bosques de coníferas del sur de California en los bosques de Baja California. De acuerdo al Inventario Forestal de Baja California 1968 (SARH 1977) las coníferas de los géneros Calocedrus, Pseudotsuga y Abies se concentran casi exclusivamente en la Sierra de San Pedro Mártir. Wiggins (1980) no hace mención de Pseudotsuga en la flora de la Baja California, como tampoco ha sido referido ni colectada por otros autores (Moran 1977, Thorne y Moran 1986, Passini et al. 1989, Peinado et al. 1994b, Delgadillo 1998). Minnich (1982) señaló que P. macrocarpa fue reportada por numerosos autores para la Sierra de San Pedro Mártir y anota que pudo existir confusión en la localidad referida. Bolton y Volgl (en Minnich 1982), sugieren que tal error podría ser debido a un «espécimen con una localidad mal interpretada». Minnich (1982) remarca que la referencia de Pseudotsuga en la Sierra de San Pedro Mártir en el Inventario Forestal de 1968 parece poco confiable por la inconsistencia de sus observaciones. Al respecto, Delgadillo (1998) sugiere que pudiera existir la posibilidad de una mala identificación y/o confusión con Abies concolor. Por otra parte, Minnich (1982) afirma que la aparente ausencia de P. macrocarpa es paralela a la ausencia o infrecuencia de otras especies de árboles acompañantes encontrados en elevaciones medias al sur de California. Y se pregunta cómo esta especie y otros elementos del bosque siempreverde declinan o desaparecen, mientras árboles mesícos del bosque de coníferas mixto continúa hacia el sur al incrementarse la aridez en Baja California. La nieve parece que juega un papel importante en los ritmos estacionales de la humedad disponible del suelo en la zona de raíces del bosque de coníferas mixto y otras comunidades de altura a lo largo de la costa del Pacífico. De hecho, aunque las altas montañas al sur de la línea internacional son enteramente mesícas, parecen deficientes para el hábitat apropiado de P. macrocarpa. 66 El bosque de coníferas Pinus juarezensis. Lanner (1974) describió en el norte de Sierra de Juárez esta nueva especie de pino piñonero, aunque con un rango de distribución hacia el sur de California y la Sierra de San Pedro Mártir (Perry 1991), creando una controversia en la aceptación de este taxa. Para Lanner, P. quadrifolia es un híbrido natural entre P. monophylla y P. juarezensis, reduciendo el estatus de P. quadrifolia como Pinus x quadrifolia. Esta especie no es referida por Wiggins (1980) en su flora de Baja California y el Manual de Jepson para la flora de California (Hickman 1993), mientras que Eguiluz (1985) la cita pero sin especificar localidad. Por su parte Farjon et al. (1997) la consideran como sinónimo de P. quadrifolia. Perry (1991) presenta una amplia descripción de P. juarezensis, quien señala además haber encontrado al oeste del poblado de La Rumorosa dos especies creciendo juntas, prefiriendo tratar a estos taxa como especies separadas (P. monophylla, P. quadrifolia, P. juarezensis). Delgadillo (1998) señala que en su trabajo de campo, y en la localidad referida, no ha sido posible localizar ningún individuo que se pudiera identificar de manera clara como P. juarezensis, debido a que todos los árboles presentaban cuatro y cinco acículas y con mayor proporción de fascículos con cuatro hojas, esto es, P. quadrifolia, requiriéndose una mayor búsqueda a detalle de esta especie. Otras especies. Wiggins (1980) hace referencia a la presencia en la Sierra de San Pedro Mártir de especies que tienen sus límites en las montañas del sur de California (por ejemplo las del Condado de San Diego). Tal es el caso de Umbellularia californica (Lauraceae), Alnus rhombifolia, Alnus tenuifolia (Betulaceae) y Arbutus menziesii (Ericaceae), sin embargo, no hay evidencias de estos taxa. El mismo autor señala que las especies de Alnus aparecen citadas en alguna literatura, y que posiblemente el fuego y el sobrepastoreo las eliminaron durante el siglo pasado. Composición florística 67 68 El bosque de coníferas Capítulo 6. Descripción de la vegetación Generalidades Dentro de las provincias florísticas Californiana-Meridional y Martirense, los bosques de coníferas se desarrollan por encima de los 1,500 msnm en las montañas de San Gabriel, San Bernandino, San Jacinto, Santa Rosa, Palomar, Cuyamaca, Laguna Juárez, y a partir de los 1,900 en la Sierra de San Pedro Mártir (Pase 1982, Peinado et al. 1994a, Delgadillo 1998). Los llamados “pinos amarillos” (Pinus jeffreyi y Pinus ponderosa) generalmente dominan estas sierras. El “pino ponderosa” es más común en la parte húmeda del Pacífico, entre 1,300 y 2,140 m; en cambio, el “pino jeffrey” es el dominante por arriba de los 1,200-1,500 m, tanto en el cismontano como en el trasmontano (Pase 1982). La vegetación de la Sierra de San Pedro Mártir es única en México, ya que representa un bosque de origen boreal muy similar en su estructura abierta y composición a los bosques de coníferas de las montañas del sur de California y, con excepción de Abies concolor, el resto de las especies de coníferas tienen su límite meridional en Baja California. De acuerdo al Inventario Nacional Forestal de Gran Visión (SARH 1992) el bosque de coníferas ocupa un superficie estatal del 2.26%. Por su parte Minnich y Franco (1998) estimaron que la superficie aproximada de estos bosques es de 335, 000 ha en las Sierra de Juárez y de San Pedro Mártir. Estos mismos autores (1997b) refieren que el bosque de coníferas en la Sierra de San Pedro Mártir cuenta 40, 360 ha, divididas con las siguientes superficies: Pinus jeffreyi, 19, 330; bosque mixto de coníferas, 15, 200; Abies concolor y Pinus lambertiana, 3,360; Calocedrus decurrens, 547; Cupressus montana 784; Pinus contorta 958; y Pinus coulteri, 154. Descripción de la vegetación 69 Resulta indudable que las coníferas, y en particular las especies dominantes del género Pinus, dan un carácter distintivo a las diferentes comunidades vegetales presentes en esta sierra. Por su carácter semiárido, estos bosques tienen un sotobosque pobre con un estrato arbustivo restringido a muy pocas especies, mientras que el componente herbáceo anual es muy diverso, tanto en primavera como en verano. Las principales especies que dominan el sotobosque son: Arctostraphylos glauca ssp. platyphylla, Ceanothus leucodermis, Symphoricarpus parishii, Salvia plachyphylla, Eriogonum wrightii ssp. oresbium, Euphorbia palmeri, Castilleja martinii, entre otras. El fuego tiene un efecto importante sobre la composición específica, la estructura y función de los bosques en la Sierra de San Pedro Mártir, y la alta frecuencia está relacionada con tormentas eléctricas, las cuales coinciden con la época de verano (Minnich et al. 1993 en Minnich y Franco-Vizcaíno 1997b). La forma en que la vegetación se quema en la sierra depende de la estructura de la misma (Minnich y Franco-Vizcaíno 1997b), y es probable que los incendios superficiales recurrentes sean la causa principal del bosque abierto en la Sierra de San Pedro Mártir, siendo estos habitualmente de intensidad moderada, con una extensión de hasta 5,000 ha y con intervalos entre incendios de aproximadamente 50 años. En los bosques mixtos de coníferas, algunos incendios pueden persistir durante varias semanas o meses. Un reclutamiento gradual de renuevos ocurre entre incendios, pero la mayoría son eliminados selectivamente por los incendios, los cuales pueden alcanzar una altura de 5 a 15 m; aparentemente existe un balance entre la tasa de ingreso y la mortalidad en las clases mayores (Minnich y Franco-Vizcaíno 1997a). Estos mismos autores refieren que, de acuerdo con sus datos, se mantiene la hipótesis de que los bosques de la Sierra de San Pedro Mártir han permanecido sin cambio durante los últimos 100 años. Dependiendo del género dominante, el bosque de coníferas de Baja California ha sido clasificado en tres tipos: bosque de Pinus, bosque de Juniperus y bosque de Cupressus (Delgadillo 1998). BOSQUE DE Pinus Rzedowski (1978) menciona que aunque la mayoría de las especies mexicanas de Pinus poseen afinidades por suelos ácidos y hacia climas templados hasta 70 El bosque de coníferas a fríos y semihumedos, existen notables diferencias entre las especies, y algunas prosperan en lugares francamente calientes, húmedos y semiáridos, así como en suelos alcalinos. Eguiluz (1985) refiere que los pinares están considerados como un género xerofítico, confinado a sitios marginados donde tienen moderada competencia con las agresivas latifoliadas. Los bosques de pinos en Baja California son una continuación de los bosques en las montañas de Cuyamaca y Palomar, en California (1,050-1,200 m), caracterizados por masas de árboles relativamente densas. De acuerdo con Eguiluz (1985) los pinares de la Rumorosa, Sierra de Juárez y Sierra de San Pedro Mártir llegaron a México a través de la Sierra Nevada. Wiggins (1980) refiere para la península de Baja California 14 especies de Pinus, y ocho de ellas se presentan en los bosques de las Sierras de Juárez y de San Pedro Mártir, y de manera exclusiva seis en esta última; las restantes especies se distribuyen en las zonas costeras e insulares. En la porción sur de la península estos bosques se presentan en la Sierra de La Laguna, por arriba de los 1,600 m, siendo Pinus cembroides y P. lagunae las especies dominantes (Delgadillo 1998). Los bosques de Pinus de las cordilleras del norte de Baja California han sido dividido en tres tipos: de montaña, costero e insular (Delgadillo 1998). En este trabajo sólo se hace la descripción del bosque de Pinus de montaña. Bosque de Pinus de montaña Estos bosques se encuentran principalmente en el norte de Baja California y en general por arriba de la vegetación de tipo chaparral, conformando el piso de vegetación a mayor altitud, entre los 1,200 y 3,000 m. Las dos grandes áreas boscosas que se localizan en el norte de la península, Sierras de Juárez y de San Pedro Mártir, presentan características especiales y diferentes en cuanto a su ecología y composición florística. Passini et al. (1989) concluyeron que los dos ecosistemas forestales son complejos y que la distribución de las formaciones de coníferas manifiesta la existencia de gradientes climáticos y altitudinales, además de una gran diferencia en sus componentes florísticos. En la Sierra de San Pedro Mártir se tienen registradas seis especies de Pinus, y su diversidad se manifiesta de manera marcada a través de un gradiente altitudinal (Delgadillo 1998). En la Sierra de Juárez, con una alti- Descripción de la vegetación 71 tud promedio altitudinal de 1,500 m, P. jeffreyi presenta una amplia dominancia, mientras que en San Pedro Mártir la comparte con otras especies de coníferas, como Pinus lambertiana, Pinus contorta var. murayana y Abies concolor. Sin duda, la Sierra de San Pedro Mártir, con promedio de 2,400 m, es donde se manifiesta la mayor presencia de especies de pináceas y de otros componentes boreales, como Cupressus montana (endémico), Calocedrus decurrens y Populus tremuloides (Delgadillo 1998). La figura 9 muestra una representación esquemática de la disposición espacial del bosque de coníferas en la Sierra de San Pedro Mártir (Delgadillo 1999). Datos del Inventario Forestal del Estado de Baja California de 1968 (SARH 1977) en la Sierra de San Pedro Mártir los pinos constituyen también el género dominante (89.6%) pero la presencia de encino es prácticamente nula (0.2%), con cierta abundancia de otras coníferas (10.2%). La poca presencia de encinos tal vez se deba a que estos árboles están por debajo de los pinos, en sitios más secos, lo que hace difícil su cuantificación. La distribución altitudinal del bosque de coníferas de la Sierra de San Pedro Mártir presentan formaciones muy definidas: a) 1,400-1,900 m, bos- FIGURA 9. REPRESENTACIÓN ESQUEMÁTICA DE LA DISTRIBUCIÓN DEL BOSQUE DE CONÍFERAS EN LA SIERRA SAN PEDRO MÁRTIR (DELGADILLO 1999). DIBUJOS TOMADOS DE ARNO (1973) 72 El bosque de coníferas ques de pinos piñonero (Pinus quadrifolia y P. monophylla); b) 1,900-2,800, bosque de P. jeffreyi; c) 2,300-3,000, bosque mixto (P. lambertiana, A. concolor y Calocedrus decurrens); y d) 2,400-2,500, bosques de P. contorta var. murrayana. Bosque de Pinus quadrifolia y Pinus monophylla En Baja California la distribución de estos pinos piñoneros van desde la frontera internacional, al norte de la Sierra de Juárez, hasta la Sierra de San Borja, particularmente el Pinus monophylla. La amplia distribucón de estos pinos en Baja California ya se conocía a fines del siglo XIX; Lemmon reportó en el Lower Californian (3 de junio de 1892) que “los bosques de piñón, compuestos de Pinus parryana o Pinus quadrifolia…. [empiezan] unas millas al sur de la frontera… y se extienden hacia el sur a lo largo de las costillas de la península, con solo unos pocos claros, hasta la punta sur de la Sierra de San Pedro Mártir”.También reportó que “otro pino piñonero, P. monophylla, mantiene una precaria existencia a lo largo de los precipios desérticos (Minnich y Franco-Vizcaíno 1999). En los escarpes orientales de toda la cordillera norte de la península la distribución de ambas especies pioneras es muy amplia. En, la Sierra de San Pedro Mártir, P. quadrifolia inicia su presencia a partir de los 1,400 m en la vertiente oeste, encontrándose la mayor densidad entre los 1,600 y 1,800 m, aunque nunca llega a formar verdaderos bosques. Sus ejemplares prefieren laderas de orientación norte, depresiones y cañadas, mezclándose con chaparral de montaña (fotos 9 y 10), comunidad esclerófila que contacta con el bosque de P. jeffreyi, por lo que algunas especies de está llegan a formar parte del estrato inferior de dicho bosque (por ejemplo, Adenostoma sparsifolium, Arctostaphylos peninsularis y Ceanothus spinosus) (Delgadillo 1999). Hacia el norte y sur de las Sierras de Juárez y de San Pedro Mártir, P. quadrifolia y P. monophylla se mezclan con el llamado chaparral desértico, mientras que las poblaciones de P. monophylla se encuentran en la vertiente oriental teniendo su límite altitudinal aproximadamente a los 1,200 m donde inicia el contacto con la vegetación micrófila desértica. También (Se halla P. monophylla en las sierras del sur del estado, como Calamujue (Yubay), La Asamblea y San Borja y San Borja (SARH 1977, J. P. Rebman, comunicación personal). Descripción de la vegetación 73 Ambas especies de pinos piñoneros se establecen en las zonas más xéricas, siendo su hábitat discontinuo y con baja frecuencia de fuegos. McCune (1988) considera a los pinos piñoneros resistentes al estrés hídrico, de escasa, altura, hojas persistentes, lento crecimiento y que requiere, varios años para producir semillas. Estos pinos crecen en hábitats fríos o secos, o ambos, y sus semillas son dispersadas por aves. Estas comunidades vegetales son muy abiertas con árboles de altura media de 8 m. La densidad registrada para P. quadrifolia en la Sierra de Juárez fue de 200 a 440 individuos /ha y una densidad media de 280 árboles /ha, con alturas de 3 a 5 m, mientras que para P. monophyla en la Sierra de San Pedro Mártir y entre los 1,400 m y 1,500 m, la densidad varió de 200 a 380 individuos /ha (Passini et al. 1989). Estas comunidades presentan tres estratos: a) arbóreo (pinos piñoneros); b) arbustivo de 1-2 m (Adensotoma fasciculatum, Adenostoma sparsifolium, Arctsotaphylos spp., Ceanothus greggii var. perplexans, Juniperus californica, Rhamanus crocea ssp. crocea, Rhus ovata, Prunus fasciculta, Quercus wislizenii); y c) suculento de 25-50 cm (Agave deserti ssp. deserti, Echinocerus engelmanii, Ferocactus acanthodes, Opuntia spp.). FOTOS 9. COMUNIDADES DE PINUS QUADRIFOLIA MEZCLADO CON CHAPARRAL, VERTIENTEOCCIDENTAL 74 El bosque de coníferas FOTOS 10. FALA DESCRIPCIÓN En las formaciones de P. quadrifolia en la Sierra de San Pedro Mártir han sido registrados 55 taxa y en las facies con Juniperus californica 42 (Delgadillo 1999). Son familias abundantes en estos bosques: Cactaceae, Fagaceae, Rosaceae, Anacardiaceae y Asteraceae (Passini et al. 1989). Pinus monophylla. McCune (1988), considera a los pinos piñoneros (P. monophylla y P. quadrifolia), de estatura corta, lento crecimiento, persistencia de sus hojas, semillas grandes (no aladas) y requiriendo varios años antes de producir semillas; la persistencia de sus hojas con un promedio siete años durante mucho tiempo puede ser considerada como una extrema expresión de plantas siempre verdes. Este mismo autor las ubica dentro de un grupo ecológico resistente al estrés hídrico, por lo que Grime (1979 en McCune 1988), lo define como «extremo contraste entre la tasa límite de producción de materia seca de toda o una parte de la vegetación». Estos pinos crecen en hábitats secos, por lo que su crecimiento lento es típico de plantas tolerantes al estrés (Grime 1979 en McCune 1988). Pinus monophylla es muy tolerante al estrés de temperatura y humedad. El retardo de la madurez sexual puede ser consecuencia del largo tiem- Descripción de la vegetación 75 po que se necesita para activar algún tamaño crítico necesario para la reproducción (Harper 1977 en McCune 1988), o de una estrategia de conservación que promueva la supervivencia individual a un nulo costo de reproducción temprana (Willson 1983 en McCune 1988). McCune (1998) considera que P. monophylla se encuentra dentro de un grupo ecológico donde se encuentran las especies de pinos vivientes más longevas (por ejemplo, P. flexilis y P. aristata). Pinus quadrifolia. El norte de Baja California es el centro de distribución de Pinus quadrifolia, ya que solamente pocas poblaciones dispersas existen al norte de la frontera internacional, en las montañas de La Laguna en el condado de San Diego, y en el monte San Jacinto. Crece en poblaciones pequeñas y compactas en medio de comunidades maduras y muy densas de chaparral de chamizo o manzanita (Minnich y Franco-Vizcaíno 1999). Ee una especie de pino que ha sido agrupado en un grupo cohesivo y distinto, resistente al estrés hídrico; es de corta altura , larga persistencia de las hojas, generalmente con grandes semillas, pero el ala corta o ausente. Corteza delgada, lento crecimiento, crece en sitios secos y requiere de muchos años antes de producir semillas. Esta especie de piñonero es de hábitats calientes y áridos, y la persistencia de las hojas es menor en comparación con P. monophylla; se distribuye en mayores altitudes hacia el norte. Al igual que otras especies de su grupo, presenta una dispersión de semillas por aves (McCune 1988). Bosque de Pinus jeffreyi Es de amplia distribución desde el suroeste de Oregón y oeste de Nevada hasta la parte norte de Baja California. Generalmente crece en altas elevaciones o en sustratos ultramáficos (Jenkinson 1980 en McCune 1988), donde está estrechamente relacionado con Pinus ponderosa, por lo que estas dos especies se traslapan y se hibridizan (Haller 1962; en Critchfield y Little 1966). Para algunos conservadores, P. jefrreyi ha sido clasificada como una variedad de Pinus ponderosa, pero ahora se acepta como una especie distinta (Farjon et al. 1977). La fragancia dulce de piña-vainilla en la resina de P. jeffreyi ayuda a determinar la especie, principalmente en días cálidos. McCune (1988) incluye a P. jeffreyi dentro de un grupo de pinos resistentes al fuego, al que sobreviven los árboles adultos; la mayoría son altos, con 76 El bosque de coníferas gruesa corteza, aciculas largas y gruesas ramas. Sus conos son pesados y con escamas que presentan espinas; semillas grandes con alas largas y son lentas para iniciar la producción de semillas. Las especies de pino varían en un orden de magnitud en edad y primera producción de semillas, en un rango de 5 a 50 años (Strauss y Ledig 1985 en Keeley y Zedler 1988). Como regla general, especies en hábitats con frecuencia de fuegos inician su reproducción más temprana que otras (Keeley y Zedler 1998). La vida media de las semillas de P. jeffreyi ha sido estimada en 120 horas en la superficie del suelo y menos del 1% no son detectadas por los predadores; en un año, más del 40% del reclutamiento de semillas y su germinación provienen de semillas ocultas y una media de distancia de 13 a 25 m de la dispersión animal (Vander Wall 1994, Keeley y Zedler 1998). Pinus jeffreyi es la especie dominante en los bosques de coníferas en el norte de Baja California. Forma grandes extensiones de masas arboladas, en un área geográficamente catalogada como semiárida y/o árida (Delgadillo 1998) (foto 11). Pase (1982) menciona que P. jeffreyi tolera un amplio rango de temperatura y humedad del suelo, pero es menos tolerante al estrés (Keeley y Zedler 1988). En la Sierra de San Pedro Mártir estos bosques son abiertos a partir de los 2,000 m, aunque llegan estar presentes hasta los 1,800 m, donde entran en contacto con el chaparral de montaña, que es su limite altitudinal inferior en los márgenes de arroyos y/o cañadas, donde se comporta como una especie riparia o freatófita. Esta distribución coincide con el límite superior de Quercus agrifolia (Delgadillo 1999) (foto 12). Passini et al. (1989) describieron este tipo de bosque como una formación con individuos mayores de más de a 15 m y una densidad que varió entre los 260 y los 540 individuos /ha y un diámetro superior a los 10 cm; la menor densidad se encontró a los 2,100 m y la mayor a los 2,400 m. Individuos con más de 25 m se registraron a los 1,700 m, al norte de la Sierra de San Pedro Mártir y representaron del 2 al 5 %. Esta comunidad presenta cuatro estratos: a) arbóreo (> 15 m); b) arbustivo (< 4 m); c) sufrutescente o subarbustivo (< 1 m), y d) el herbáceo (< 50 cm). Se han registrado 106 táxones y esta riqueza florística se debe a la gran extensión altitudinal que cubre la sierra (Passini et al. 1989). Descripción de la vegetación 77 FOTO 11. PANORÁMICA HACIA EL OESTE DEL BOSQUE DE PINUS JEFFREYI (2700 M) FOTO 12. PINUS JEFFREYI Y QUERCUS AGRIFOLIA A LO LARGO DE CAÑADAS Y DEPRESIONES, RODEADOS DE CHAPARRAL (1900 M) 78 El bosque de coníferas Bosque mixto Formación boscosa característica de la parte alta de la sierra, que tiene su crecimiento óptimo entre los 2,300 –2,800 m. Passini et al. (1989) la refieren como una faciación de P. jeffreyi y como una formación con P. jeffreyi, P. lambertiana y A. concolor como co-dominantes; esta última especie prefiere laderas sombreadas (exposición norte). Hacia el sur y por encima de los 2,300 m, Calocedrus decurrens es común pero no abundante, principalmente en la zona de La Tasajera (Delgadillo 1999). Este tipo de bosque muestra una disposición espacial abierta, con un sotobosque escaso, y con cuatro estratos: a) arbóreo (> de 40 m); b) arbustivo (1-4 m); c) sufrutescente o subarbustivo (< 50 cm); y d) y herbáceo (< 30 cm). Las facies de A. concolor, con o sin P. lambertiana, presentan una densidad entre 140 y 380 individuos/ha, se consideran individuos con más de 8 m de altura (diámetro superior de 10 cm); las coberturas son de 89% a 53% de P. jeffreyi, 47% a 11% de A. concolor y P. lambertiana no pasa del 15%. La superficie del terreno de las facies varia de 17 a 44 m²/ha, con un valor medio de 29 m²/ha. La densidad de la formación de A. concolor como dominante, comprende de 220 a 440 individuos/ha (diámetros superiores a 10 cm), y constituye del 95% al 55 %, con un valor medio de 81% y P. jeffreyi de 5 a 25%; P. lambertiana y C. decurrens no representan más del 9% del total de individuos. La superficie del terreno comprendió entre 48 y 61 m²/ha y un valor medio de 51 m²/ha (Passini et al. 1989). Pinus lambertiana. Especie dentro de los llamados “pinos blancos” se distribuyen en el oeste de Oregón, las cordilleras costeras de California, la Sierra Nevada y las montañas del sur de California y Nevada, extendiendo su presencia en la Sierra de San Pedro Mártir (Critchfield y Little 1966). Se encuentra entre los 2,000 y los 3,000 m de elevación (Perry 1991), con divergencia y probable afinidad filogenética con especies asiáticas (McCune 1988). Esta especie es característica del componente montano con un rango considerable en latitud, pero rara vez en dominante (Keeley y Zedler 1998). Representan a los árboles de pinos más altos de Norteamérica, de más de 60 m y diámetros mayores de 2 m. Crecen en sitios húmedos, suelos arenosos y gravosos bien drenados, teniendo un mejor crecimiento en hondadas y sitios fríos (Perry 1991). Son árboles que pueden alcanzar gran Descripción de la vegetación 79 longevidad (Fowell 1965 en Keeley y Zedler 1998), y debido a su gran tamaño los árboles más viejos pueden tener cierta resistencia, por lo que su distribución se sobrepone con especies de mayor tolerancia al fuego como P. jeffreyi y P. ponderosa (Keeley y Zedler 1998). Su tolerancia a la sombra también permite el establecimiento de árboles jóvenes por debajo del dosel, en pequeños huecos (Keeley y Zedler 1998). Pueden germinar en suelo mineral o litter (Rundel et al. 1977, en Keeley y Zedler 1998) y retrasar la germinación de sus semillas durante muchas décadas (Keeley y Zedler 1998). McCune (1988) incluye a P. lambertiana en el mismo grupo de pinos resistentes al fuego, donde se encuentra P. jeffreyi. Como regla general, las especies en hábitats con alta frecuencia de fuego inician su reproducción más temprana que otras, pero algunas especies que viven en ambientes moderadamente productivos, como P. lambertiana, también retrasan su reproducción durante varias décadas (Keeley y Zedler 1998). Sin embargo, P. lambertiana puede invadir áreas recientemente quemadas (Talley y Griffin 1980 en Keeley y Zedler 1998). Pinus coulteri. Especie restringida a las montañas de la costa y a la parte central y sur de California y norte de Baja California. En zonas de traslape está relacionada con Pinus jeffreyi y Pinus sabiniana, y es conocida la hibridación en la naturaleza con Pinus jeffreyi (Zobel 1951 en Critchfield y Little 1966). En Baja California, P. coulteri se distribuye de manera dispersa y rara en la Sierra Blanca (1,200 m), Sierra de Juárez (1,200-1,800 msnm) y Sierra de San Pedro Mártir (1,900-2,150 msnm). En las Sierras de Juárez y de San Pedro Mártir, P. coulteri se adapta mejor a los de roca granítica de suelos someros, mientras que P. jeffreyi a suelos tanto profundos como someros. Esta especie de pino que forma parte de un grupo resistente al fuego, al que sobreviven los árboles adultos; la mayoría son altos, con gruesa corteza, aciculas largas y gruesas ramas. Sus conos son pesados y con escamas que presentan espinas; semillas grandes con alas largas, y son lentas para iniciar la producción de semillas. Es menos divergente del grupo (P. sabiniana, P. torreyana), con lento crecimiento y de tamaño menor; la parcialidad serotina aparentemente le da una ventaja reproductiva después del fuego sobre otras especies del grupo (P. lambertiana) (McCune 1988). P. coulteri, la igual que P. torreyana (Bradron 1949; Vogl 1973 en Keeley y Zedler 1998), tiene conos que mantienen el árbol por años y se abren al ma- 80 El bosque de coníferas durar sin estimulo del calor. Una gran porción de las semillas salen en las primeras semanas, pero algunas son mantenidas en la porción proximal del cono y caen durante grandes periodos, posiblemente en respuesta al movimiento gradual causado por la humedad y la sequía (Keelley y Zedler 1998). Especie ecológicamente muy importante, se distribuye en el norte de Baja California en reducidas extensiones, principalmente en Sierra Blanca (1,200 m), Cerro Hanson, en la Sierra de Juárez (1,600 m) (Delgadillo 1988), así como al norte de la Sierra de San Pedro Mártir, entre los 1,900-2,150 m (Minnich 1986). Aunque P. coulteri y P. lambertiana han sido colocados en el mismo grupo por McCune (1988), la aparente serotinidad parcial de la primera especie le confiere una ventaja reproductiva sobre la segunda (Griffin 1982 en McCune 1988). Por lo general, las poblaciones de P. coulteri se desarrollan en contacto con el chaparral, y están sujetas a una dinámica del fuego (Delgadillo 1998), sin embargo, las relaciones entre P. coulteri y el fuego no están muy clara (Vale 1979). Abies concolor. Especie de abeto que se distribuye desde Oregón, en las montañas de Siskiyou y Cascadas, Sierra Nevada hasta el sur de California y norte de Baja California. También se encuentra en algunas partes al oeste de Idaho, Wyoming y sur de Colorado, sur de las Montañas Rocosas de Utah y Colorado, sur de Arizona, Nuevo México y así com norte de México (Sonora y Chihuahua). De todos los abetos del oeste, A. concolor es el de mayor distribución, y es reconocido por su alta productividad. El máximo tamaño y mejor desarrollo se encuentra en la parte central de Sierra Nevada, en California, donde el récord de especímenes es de 58.5 m de altura y 271 cm en la base. Es generalmente , es tolerante a una variedad de condiciones del suelos, disponibilidad de nutrientes y pH. Es más dependiente a la disponibilidad de humedad y la temperatura que a los suelos, siendo moderadamente sensitivo a los excesos de humedad en el suelo. Estudios en California y Oregón sobre la producción de conos, indican que tienen un ciclo de 3 a 9 años en condiciones duras; buenas producciones se presentan generalmente de 2 a 5 años. En sitios extremos, la producción de conos puede presentar patrones variables (Laacke 2004). El tamaño de las semillas es variable, y un kilo puede contener entre 18,960 y 39,070.Porciones relativamente pequeñas de las semillas están Descripción de la vegetación 81 sanas (20-50%), inclusive en un año bueno. La producción de semillas varía con la edad del árbol, tamaño y dominancia; las semillas germinan en la primavera, después de derretirse la nieve. En general, los abetos blancos se establecen al germinar en la sombra parcial, pero una vez allí crecen favorablemente con luz completa. Es menos tolerante a la sombra que otros abetos verdaderos (Laacke 2004). El único abeto peninsular se encuentra en la Sierra de San Pedro Mártir por arriba de los 2,200 m, bajo condiciones de una alta humedad atmosférica y edáfica y prefiere las laderas de orientación norte (ombría) donde forma grandes poblaciones (Delgadillo 1999). Al respecto Pase (1982) refiere que la exposición norte y este, la humedad elevada y las elevaciones mayores, dan como resultado el incremento de la abundancia A. concolor y la disminución de P. jeffreyi y P. lambertiana, asociación boreal que incluye a Calocedrus decurrens (Delgadillo 1998). Bosques de Pinus contorta var. murrayana Pinus contorta se distribuye desde la parte central de Yukon, en el oeste de Canadá, hasta el Distrito de Mackenzie, en las Montañas Liard, así como el distrito suroeste de Colorado y en la parte norte de Baja California. Es común observarlo a lo largo de la costa del Pacifico a una altitud de aproximadamente los 3,300 m al sur de la Sierra Nevada y las Montañas Rocosas. La diversidad morfológica y su amplio rango de distribución ha sido estudiada por Critchfield (1957 en Critchfield y Little 1966). Muchos especialistas reconocen dos formas: la primera, arbustos pequeños que crecen en las costas del Pacifico, y la segunda, con mayor altura, que crece tierra adentro en altitudes de 1,500 a 3,300 m. Prefiere sitios húmedos, tanto costeros como de alta montaña (McCune 1988), Especie de conos serotinos pequeños y abiertos, polimórficos y con poblaciones o taxa subespecificos de cono no cerrados, o poblaciones polimórfitas, o individuos con ambos tipos de cono (cerrados y abiertos). Las altas intensidades de los fuegos pueden seleccionar la dureza, asimetría, lo sésil de los conos, y pueden reducir los cambios de ignición con una mejor protección para las semillas. Aunque tiene baja tolerancia al fuego, responde a él produciendo abundantes semillas de tamaño pequeño, de liberación retardada y germinación precoz. Son individuos de baja altura, 82 El bosque de coníferas hojas cortas y persistentes, cuyos conos no son generalmente muy serótinos (McCune 1988). La variedad murrayana es un pino subalpino en ambientes que se caracterizan por una etapa corta de crecimiento, baja acumulación de materia orgánica, infrecuentes periodos de sequía y baja incidencia de fuegos (Parker 1986, Sheppard y Lassoie 1985 en Keeley y Zedler 1998). En Baja California las poblaciones de P. contorta var. murrayana sólo se encuentran en la Sierra de San Pedro Mártir y tienen su óptimo de crecimiento en los márgenes de las praderas de Vallecitos (2,450 m) y cerca de arroyos someros, siendo freatófitas obligadas por sus altos requerimiento se agua. Es una formación dominante con árboles de 10 a 15 m a la cual, algunas veces lo acompaña P. jeffreyi (Delgadillo 1999). La densidad registrada es de 140 a 320 individuos/ha y diámetro superior a 10 cm; su porcentaje es de 100 a 60%, con un valor medio de 68%. La superficie media del terreno es de 20 m²/ha y un valor máximo de 38 m²/ha (Passini et al. 1989). Bosque de Juniperus Miranda y Hernández X. (1963 en Rzedowski 1978) incluyen a este tipo de comunidad vegetal en la categoría de «bosques de escuamifolios». Rzedowski (1978) refiere que cubre grandes áreas en México y se halla bien distribuido desde Baja California y Tamaulipas hasta Chiapas, prosperando en diversas condiciones ecológicas. La superficie nacional cubierta por este tipo de bosques es menor al 0.04% (Flores et al. 1971). Se le encuentra preferentemente en forma de una estrecha faja transicional entre el bosque de Quercus y Pinus por un lado, y el pastizal, matorral xerófilo o bosque tropical caducifolio, por el otro. Las áreas arboladas de la asociación siempreverde de Juniperus y pino piñonero tuvieron su centro evolutivo en la Gran Cuenca, y es uno de los tipos de vegetación más extendidos en el suroeste de los Estados Unidos. Abarca hacia el sur desde Colorado, Utah, Nevada, norte y centro de Arizona, sur de Nuevo México a las áreas montañosas de Transpecos (Texas), sur de California y norte de Baja California. Esta asociación cubre extensas áreas entre los 1,000 y 2,300 m (extremos de 1,050 y 2,700), y tienen su desarrollo en mesetas, mesas, llanuras, laderas y cordilleras (Brown 1982). Con frecuen- Descripción de la vegetación 83 cia los bosques de Juniperus no parecen constituir una comunidad clímax, sino de origen secundario, aunque este aspecto no ha sido estudiado a fondo en ninguna parte de México. Estos bosques, especialmente en el centro de país, se ven muy perturbados por el pastoreo y por la tala, y posiblemente representan fases de sucesión secundaria, cuyo clímax corresponde al pinar o encinar. Los nombres comunes más frecuentes que reciben las especies de Juniperus en México son cedro, sabino, enebro, nebrito, tlaxcal y táscate (Rzedowski 1978), y huata en Baja California (Delgadillo 1998). Muy pocas veces Juniperus y Cupressus han sido descritos como un componente principal de las áreas boscosas de Baja California, ya que a diferencia de los bosques de pino, no ocupan grandes extensiones; sin embargo, su papel florístico y ecológico dentro del mosaico de la vegetación es muy importante. En Baja California se encuentra una sola especie, Juniperus californica, que se distribuye a manera de “parches” (por ejempl, en la Sierra de Juárez, Sierra de San Pedro Mártir y las islas de Cedros y Guadalupe) (Delgadillo 1998). Las poblaciones peninsulares más sureñas se encuentran formando reliquias en las sierras de Calamujué y San Borja, en altitudes que van desde los 800 a los 1,700 m en la parte peninsular, mientras que en isla de Cedros se presenta entre 75-1,200 m, y en isla de Guadalupe entre los 15-350 m (Zañoni y Adams 1973). Por lo general, se encuentra en hábitats secos y asociada a chaparral de montaña y bosques de pino piñonero (P. monophylla y P. quadrifolia), aunque algunas veces esta especie llega a encontrase de una manera pura. Se desarrolla en diferentes tipos de suelos, desde los profundos y graníticos hasta los pedregosos y en ocasiones sobre calizas (Delgadillo 1998). La distribución de J. californica en la Sierra de San Pedro Mártir, se presenta en el norte y oeste, entre los 700 y 1,500 m. Su forma es arbustiva de hasta cuatro metros, y es resultado de un patrón evolutivo de convergencia propio de la vegetación mediterránea (chaparral) (Delgadillo 1999). Las relaciones y sinonimias de J. californica no han sido tratados con detalle en México (Zañoni y Adams 1973), en Estados Unidos se relaciona de manera cercana con J. monosperma, que se distribuye al oeste de Texas, centro y sur de Nuevo México, Arizona y sur de California (Brown,1982). Vasek (1966 en Zañoni y Adams, 1973) discutió la distinción entre J. californica y sus relaciones con J. occidentalis y J. osteosperma, por lo que J. 84 El bosque de coníferas californica la sido considerada como sinónima de J. occidentalis por varios autores europeos. Al igual que otras coníferas en Baja California, la aridez es el factor que limita el crecimiento de Juniperus californica. Al germinar puede llegar hasta 30 cm, por lo que depende mucho de la sombra, debido posiblemente a que pueden reemplazar a los juníperos que están creciendo; su crecimiento es encorvado (torcido), bifurcado y con multitallos sus ramas son tiesas con tallos irregulares. Al madurar el árbol puede alcanzar de 1 a 4.5 m, y en ocasiones llega a los 12 m. Cada gálbulo contiene dos semillas que son dispersas por aves y mamíferos, que se comen la parte carnosa del gálbulo, escarificando las semillas. No se cuenta con reportes acerca del mínimo de edad de las semillas. Las estructuras sexuales aparecen en dicha estación y la germinación a finales también de dicha estación. En ausencia de alteración (fuego u otros), se puede desplazar en áreas con árboles maduros o muertos. El agua es un factor que limita su crecimiento; la edad de los árboles no es la mayor influencia en las tasas de crecimiento véase: (http:/referente.allrefer.com/wildlife-plants-animals/ plants/tree/). Bosque de Cupressus Este tipo de bosque, considerado como reliquia, está restringido a ciertas áreas con ambientes ecológicos muy específicos, que le permiten desarrollarse vigorosamente, aunque con limitantes en la regeneración de su población. Esta comunidad vegetal también forma parte de los «bosques escuamifolios» de Miranda y Hernández X. (1963 en Rzdedowski 1978). Las especies de Cupressus en muy pocos lugares llegan a ser dominantes en la vegetación de México, aunque en algunas regiones convive con Abies, y a veces con Pinus y Quercus. Este género está restringido a ciertas condiciones costeras (Brown 1982). Estos bosques fueron más extensos antes del Pleistoceno (Mason 1932; Axelrod 1967 en Brown 1982). Tres especies aparecen exclusivamente en el noroeste de la península: C. arizonica, C. montana y C. forbesii, y C. guadalupensis ssp. guadalupensis en Isla de Guadalupe (Delgadillo 1998). Cupressus montana. Es una especie endémica de la Sierra de San Pedro Mártir, distribuyéndose por arriba de los 2,300 msnm, es un árbol bajo de Descripción de la vegetación 85 hasta siete m y de forma cónica. Goldman (1916) la registró como C. guadalupensis en su expedición a Baja California. Este ciprés está asociado principalmente con Abies concolor, Pinus lambertiana, P. jeffreyi, Calocedrus decurrens y Populus tremuloides, y tiene su mejor desarrollo en cañadas al sur y este de la Sierra (La Tasajera, La Grulla y Bajío de las Viejas) (Delgadillo 1998). Calocedrus decurrens. Representa al tercer género de la familia Cupressaceae en Baja California. Se distribuye a lo largo de la Sierras Nevada y Rocosas, en bosques mixtos y asociado a P. lambertiana, Abies concolor y Pseudostuga macrocarpa entre los 1,700 y 2,600 m, y por debajo de las series de bosques de P. jeffreyi y P. ponderosa. Las poblaciones de C. decurrens en Baja California se encuentran principalmente al sur de la Sierra de San Pedro Mártir, pero sin formar manchones puros al igual que en el sur de California; se mezcla con P. lambertiana, A. concolor, P. jeffreyi y Populus tremuloides. Su hábitat son las laderas, sitios protegidos y próximos a los arroyos. Los individuos alcanzan hasta 40 m y diámetros de cerca de dos m. Es muy común encontrar árboles adultos con ápices del tallo roto debido a nevadas y fuertes vientos en invierno, además de relámpagos en verano. Se han encontrado algunos ejemplares en Sierra de Juárez, y que probablemente han escapado de algunas pequeñas plantaciones de ranchos y del vivero forestal dentro del Parque Nacional Constitución de 1857 (Delgadillo 1998). Vegetación riparia Dentro del bosque de coníferas de la Sierra de San Pedro Mártir se encuentran comunidades riparias, que forman una estrecha interfase entre los ecosistemas acuáticos y terrestres en las regiones montañosas del noroeste de la costa del Pacífico de Norteamérica (Youngblood et al. 1985 en Kovalckik y Chitwood 1990). Aunque la península de Baja California no se caracteriza por la abundancia de este tipo de corrientes de agua, la región noroeste, incluídas sus sierras, cuentan con algunos arroyos de regular caudal en la vertiente del Pacífico, originados principalmente en las montañas costeras y en las sierras de Juárez y de San Pedro Mártir (Delgadillo 1998). Los ambientes riparios están protegidos de fuertes vientos y de los veranos secos extremosos. Las inundaciones causan destrucción de la vege- 86 El bosque de coníferas tación y crean nuevos sitios para el establecimiento y regeneración de nuevas especies (Triska et al. 1982 en Swason et al. 1982). Towe (1964) presenta la definición más sucinta y ampliamente aceptada de las comunidades riparias al establecer esta se presenta en el área adyacente al canal de agua y/o en las zonas de inundación, y que se caracterizan por especies y/o formas de vida diferentes a los de clímax no riparios que las rodean. Durante muchos años los ecólogos se han referido a los taxa riparios caducifolios de hoja ancha como especies riparias obligadas: Populus, Salix, Fraxinus, Junglans, Platanus y otros. Este y otros términos, como riparios facultativos y pseudorriparios, han sido aplicados a diversos taxa “riparios”. El Servicio de Pesca y Vida Silvestre de los EE UU. adoptó la siguiente nomenclatura: ripario obligado por el “ripario verdadero” y ripario facultativo por el de “pseudorripario” (Reichenbacher 1984). Rzedowski (1994) refiere que Platanus, Populus y Salix son de franca extracción holártica. Algunos de estos últimos géneros descienden altitudinalmente a lo largo de los ríos hasta áreas de clima caliente, no pocas veces cercanas al nivel del mar, donde las galerías de Fraxinus, Platanus, Populus y Salix están flanqueadas por una vegetación termofila, integrada por elementos de vinculación tropical. La presencia de estas poblaciones constituye una situación relictual de épocas más frías del Cuaternario y del Terciario. Se ha reconocido la asociación Populus-Salix como una colonizadora agresiva en sitios ecológicamente alterados (Reichenbacher 1984). La distribución actual de las especies de Populus es el resultado de una larga secuencia de desplazamientos geográficos que incluyen tanto invasiones como regresiones, y seguramente extinciones (Rzedowski 1985). Especies como Platanus racemosa, Populus fremontii y Salix lasiolepis han sido registradas en el sur de EE UU. donde aparentemente contribuyeron a una vegetación arbustiva subárida (geoflora del madro-terciario), que se extendió fuera de esta región con expansión en el clima seco del Terciario medio y tardío (Axelrod 1958 en Robichaux 1977). Originalmente estas comunidades riparias ocuparon muchas de las áreas de escurrimiento en las provincias bióticas Californiana, Mojaviana, Sub-Mogollona (=arizonica), Madreana y Chihuahuana, pero actualmente se han visto reducidas por la disminución de las corrientes (Minckey y Brown 1982). Tres estratos componen estas comunidades: arbóreo, arbustivo y herbáceo, además de plantas acuáticas. El componente florístico de las zonas Descripción de la vegetación 87 riparias de Baja California se caracteriza por tener árboles caducifolios en la época invernal, que algunas veces llegan a medir hasta 15 m, siendo los géneros Platanus, Salix y Populus los únicos en el estrato arbóreo. A veces se presenta Quercus agrifolia (encino), pero este con un comportamiento más de tipo freatofito (Delgadillo 1999). Las comunidades riparias que se encuentran en el bosque de coníferas de la Sierra de San Pedro Mártir tienen como especies características a Populus tremuloides (alamillo) y Salix lasiolepis (sauce). Poblaciones de Populus tremuloides (secc. Populus) (foto 13), se presentan en casi todos los arroyos del bosque de coníferas, siendo una especie ampliamente extendida en Norteamérica y ligada a climas fríos y en muchas partes llega hasta los límites con la tundra. En México se le encuentra con cierta frecuencia en el norte de la Sierra Madre Occidental, pero en otras regiones su presencia es esporádica y aislada (Rzedowski 1985). Según Minnich y Franco-Vizcaíno (1999) Populus trichocarpa se presentan en la Sierra de San Pedro Mártir sobre el arroyo de La Grulla y el río San Rafael. En contraste, Salix lasiolepis aparentemente no sólo sobrevivió sin modificaciones en el sudoeste y California, sino también extendió su distribución hacia el norte, hasta la Columbia Británica. Esto ocurrió sólo en las regiones interiores secas del norte y no a lo largo de la franja costera, lo que refleja sus orígenes sureños (Robichaux 1977). Algunos de los principales taxa acuáticos o en márgenes de los arroyos, que se encuentran en las zonas riparias del bosque de coníferas son: Berula erecta, Carex spp., Ceratophyllum demersum, Eleocharis spp., Juncus spp., Lemna minor, Lemna trisulca, Mimulus spp., Potamogeton pectinatus, Potamogeton natans, Ranunculus cymbalaria, Rorippa nasturtium-aquaticum, Scirpus americanus, Scirpus koilolepis, Urtica dioica, Zannichelia palustris y Zauschneria californica. Praderas Dentro de este bosque encontramos tambien praderas de montaña con presencia de especies herbáceas, en su mayoria perennes, que representan zonas ricas en alimento e indispensables para la fauna silvestre. Sin embargo, estos prados han sido utilizados de manera intensiva para el pastoreo de ganado bovino, principalmente durante los veranos. Estas 88 El bosque de coníferas zonas de praderas se encuentran en Vallecitos, La Grulla, La Encantada y Santa Rosa. Algunas de las especies más importantes son: Potentila wheeleri, Trifolium wigginsii (endémica), Achillea millefolium, Eriogonum hastatum, Linanthis melingii, Muhlenbergia richardsonis, Ophiocephalus angustifolius (endémica) Lesquerella peninsulares (endémica), y especies de los géneros Juncus, Scirpus y Eliocharis. FOTO 13. COMUNIDAD DE POPULUS TREMULOIDES (ÁLAMO), CERCA DE VALLECITOS. SIERRA DE SAN PEDRO MÁRTIR Descripción de la vegetación 89 Capítulo 7. Comunidades fitosociológicas Generalidades La fitosociología sigmatista o de Zurich-Montpellier está basada en una metodología de análisis de la vegetación que se aplica de manera muy amplia en todo el mundo, fue desarrollada a principios del siglo XX en países europeos cuya lengua no era el inglés como una alternativa de clasificación de la vegetación en base a la fisionomía (con la formación como unidad base) (Jaroslav 1992). Este método consiste básicamente en la toma de inventarios (muestreos) en comunidades homogéneas climáticamente y la asignación de un valor numérico (uno al cinco) años a los taxa encontrados para referir un índice de abundancia/cobertura, siendo 5 años el valor más alto (Braun-Blanquet (1932 en Dansereau 1957) (cuadro 9). A partir de estos inventarios, se podrán reconocer aquellas especies con valores constantes y de alto porcentaje de frecuencia. La finalidad de la fitosociología es conocer la unidad básica: la asociación vegetal, una comunidad vegetal de carácter abstracto, a cuyo conocimiento se llega mediante el estudio comparativo de la composición florística. El término asociación es usado en la fitosociología en el sentido dado por Braun-Blanquet (1932 en Dansereau 1957) que denota una comunidad de plantas de cualquier rango, ya sea efímera pionera o una climácica perdurable que posee cualidades florística, ecológicas, biogeográficas, dinámicas, catenales e históricas pecualiares. Una nomenclatura binomial, similar a la Linneo, se utiliza para designar las asociaciones. El nombre de la asociación está formado a partir del (los) Descripción de la vegetación 91 CUADRO 11. ÍNDICE DE ABUNDANCIA-COBERTURA DE BRAUN-BLANQUET (1932; EN DANSEREAU 1957). 5 4 3 2 1 + Cualquier número de individuos, pero cubriendo más de tres cuartas partes de la superficie. Cualquier número de individuos, pero cubriendo entre un la mita y tres cuartas partes de la superficie. Cualquier número de individuos, pero cubriendo entre un cuarto y la mitad de la superficie. Individuos muy abundantes con cobertura escasa o bien cubriendo menos de un cuarto de la superficie. Individuos bastante abundantes pero con baja cobertura. Planta escasa con un valor de cobertura pequeña. Fuente: Braun-Blanquet 1932 en Dansereau 1957. nombre(s) científico(s) válidamente publicado(s) de una o dos especies, o de táxones infraespecíficos, que forman parte de la diagnosis original, a lo que se añade la desinencia de rango al radical del género; así la desinencia –etum se agrega al género de la especie dominante (artículo 10 del Código de Nomenclatura Fitosociológica 1988, Backman et al. 1988). Las asociaciones se reúnen en jerarquías o rangos principales de orden creciente, aplicándose la desinencia de la siguiente manera: clases, –etea; ordenes, -etalia; y alianzas, -ion (Rivas-Martínez 1987). Algunos ecólogos estadounidenses han puesto de relieve la necesidad de aplicar la metodología fitosociológica para la clasificación de las comunidades vegetales norteamericanas (Major y Taylor 1988 Rundel y Vankat 1989). Sin embargo, y por diferentes razones, esta metodología no ha sido desarrollada de manera amplia en Norteamérica, a excepción hecha de Canadá (Delgadillo 1995). A continuación se describen las comunidades vegetales reconocidas fitosociológicamente hasta la fecha en el bosque de coníferas de la Sierra de San Pedro Mártir, y de acuerdo a las siguientes jerarquías (esquema sintaxonómico) (Peinado et al. 1994a, 1994b, 1995, 1997, Delgadillo 1995). Clase Ephedro viridis- Juniperetea osteospermae Orden Rhoo anisophyllae-Pinetelia monophyllae 92 El bosque de coníferas Alianza Rhoo anisophyllae-Pinetiona monophyllae Asociación. Junipero californicae- Pinetum monophyllae Asociación. Adenostomo sparsifolii- Pinetum quadrifoliae Clase Arctostaphylo patulaee-Pinetea lambertianae Orden Amelanchiero pallidae-Pinetalia jeffreyi Alianza Pinus jeffreyi-Abietion concoloris Asociación. Symphoricarpo oreophili-Pinetum jeffreyi Orden Polygono davisiae-Pinetalia murrayanae Alianza Polygono davisiae-Pinion murrayanae Asociación. Potentillo rimicolae-Pinteum murrayanae Bosques de pino piñonero y juniperus CLASE FITOSOCIOLÓGICA: EPHEDRO VIRIDIS-JUNIPERETEA OSTEOSPERMAE Vegetación forestal abierta dominada por sabinas (Juniperus sect. sabina) y pinos piñoneros (Pinus edulis, P. monophylla, P. quadrifolia), adaptados a climas muy continentales y en ombroclimas semiárido y seco. Estos bosques aparecen en la literatura inglesa como «Pinyon-Juniper Woodland» (Barbour 1988, West 1988, Brown 1982, Axelrod 1988, Vasek y Thorne 1988), Los actuales bosques de piñoneros se extienden en fundamentalmente en las zonas de altitud media situadas entre el eje Cascadas-Sierra Nevada y las Rocosas, aproximadamente entre los 45º y 37º N (Axelrod 1958, 1975, 1976, 1979, 1981, 1988 Raven y Axelrod 1985, Raven 1988, Wells 1983, Betancourt et al. 1990 Barbour y Christensen 1993). Estos bosques de piñoneros y sabinas, que se extienden básicamente desde las montañas de Santa Catalina y Chiricahuas en Arizona meridional, hasta el Transpecos y la cuenca del río Grande en Texas, se caracterizan tanto por sabinas (Juniperus ashoi, J. erythrocarpa, J. pinchotii, J. deppeana) y pinos piñoneros (Pinus cembroides var. remorata, P. cembroides var. bicolor) así como por encinos endémicos. Por el contrario, las comunidades de piñoneros situadas más al occidente constituyen una manifestación empobrecida del antiguo bosque Madreano, por haber quedado en las zonas mas favorables tanto térmica como ómbricamente. Su extensión superficial, de acuerdo con las estimaciones de West (1988), es de unos 17 millones de hectáreas sólo en los Estados Unidos. En función del régimen de Descripción de la vegetación 93 precipitaciones pueden distinguirse dos grandes tipos de bosques de sabinas y piñoneros, a los que hemos dado la categoría de orden. ASOCIACIÓN JUNIPERO CALIFORNICAE-PINETUM MONOPHYLLAE (PEINADO ET AL. 1994B) CARACTERÍSTICA TERRITORIAL: PINUS MONOPHYLLA Estructura. Bosque abierto de mediana altura (4-6 m), donde la especie dominante es el pino piñonero (P. monophylla), que raramente juntan las copas, por lo que quedan numerosos espacios abiertos que son ocupados por sabinas arbustivas (rara vez por arbolillos con troncos definidos) de 1-3 m de altas; el resto son un cortejo muy heterogéneo de arbustos y cactáceas, variable en su composición de acuerdo con el área geográfica. Además de la estructura típica de la asociación, se han distinguido dos variantes: a) de Quercus berberidofolia, existente en algunos valles en sombra de lluvia del sector Venturano (provincia Californiana-Meridional); y b) carente de P. monophylla, que parece coincidir con áreas quemadas y una constancia de Yucca schidigera y Adenostoma fasciculatum que sugiere una evolución posterior al fuego a partir de la asociación Yucco schidigeraeAdenostometum fasciculati (Peinado et al. 1997; Delgadillo 1998). Ecología y fitogeografía. Asociación mesomediterránea semiárida, existente en las provincias Californiano-Meridional y Martirense, así como en algunas montañas de la provincia Colorada (vertientes meridionales del Joshua Tree National Monument). Sinfitosociología. Etapa madura de la serie mesomediterránea semiárida Californiano-Meridional, Martirense y Colorada del pino piñonero Pinus monophylla (Junipero californicae-Pineto monophyllae S.). ASOCIACIÓN ADENOSTOMO SPARSIFOLII-PINETUM QUADRIFOLIAE PEINADO ET AL. 1994B Estructura. En condiciones óptimas se trata de un bosque abierto, fácilmente transitable, dominado por Pinus quadrifolia (5-10 m de alto), que constituye un mosaico de árboles que rara vez llegan a juntar las copas. En consecuencia, en los abundantes claros del bosque coexisten, además de Adenostoma sparsifolium y Juniperus californica, un conjunto heterogéneo de táxones heliófilos formado tanto por esclerófilos (Rhus ovata, 94 El bosque de coníferas Arctostaphylos spp., Quercus chrysolepis, Garrya grisea, Prunus ilicifolia), como por elementos de origen neotropical (Opuntia, Agave, Ferocactus,Yucca). Este parece ser el estado original de la comunidad, cuya estructura es coincidente con otros bosques de pinos de cono cerrado (por ejemplo, P. monophylla), con los que están ecológica y florísticamente relacionados. Sin embargo, por acción del fuego que ha castigado fuertemente a este tipo de bosques fácilmente incendiables (Minnich 1983), la comunidad evoluciona hacia un aspecto dominado por Adenostoma sparsifolium, donde los pinos quedan como ejemplares aislados o llegan a faltar por completo. En el límite altitudinal de P. quadrifolia en la Sierra de San Pedro Mártir el pinar se enriquece con elementos supramediterráneos como Pinus jeffreyi, Arctostaphylos patula var. platyphylla, entre otras especies. Ecología. Asociación mesomediterránea que prospera bajo ombroclimas que oscilan desde el semiárido superior al seco. Conforme deciende la precipitación, lo que ocurre en las vertientes orientales de las Sierras de Baja California, estos pinares son sustituidos por Junipero californicae-Pinetum monophyllae. Fitogeografía. Asociación cuyo óptimo se encuentra en las montañas del noroeste de Baja California (Sierras de Juárez y de San Pedro Mártir), pertenecientes a la provincia fitogeográfica Martirense. De forma relictual, la asociación se encuentra también en las vertientes más áridas de algunas montañas de la provincia Californiano-Meridional (San Jacinto, Santa Rosa y Laguna). Sinfitosociología. Es la etapa madura de la serie silicícola mesomediterránea semiárida seca, Martirense y Californiano-Meridional de Pinus quadrifolia (Adenostomo sparsifolii-Pineto quadrifoliae). BOSQUE DE Pinus de MONTAÑA CLASE FITOSOCIOLÓGICA: ARCTOSTAPHYLO PATULAE-PINETEA LAMBERTIANAE Bosques de P. jeffreyi y mixtos propios de los pisos supramediterráneo y oromediterráneo de las altas montañas de la región Californiana. Por lo general, prosperan bajo ombroclima al menos seco; por el descenso de las precipitaciones tanto en el piso supramediterráneo como mesomediterráneo, son sustituidos por bosques abiertos de sabinas y pinos piñoneros Descripción de la vegetación 95 (Ephedro viridis-Juniperetea osteospermae), mientras que en el piso mesomediterráneo bajo ombroclima seco o subhúmedo son reemplazados por encinares de la clase Heteromelo arbutifoliae-Quercetea wislizeni. Barbour y Christensen (1993) mencionan que este tipo de bosque, incluido en los denominados «Western Montane Coniferous Forests», es un amplio grupo muy heterogéneo desde el punto de vista florístico y ecológico, que incluye todos los bosques de coníferas, exceptuando los bosques de coníferas del Pacífico situados al occidente de las Rocosas, desde los 65° a los 19° N y los 100° a los 145° de longitud oeste. El soporte florístico que sustenta este tipo de bosques montanos del oeste es muy bajo; entre los árboles tan sólo Populus tremuloides, Pseudotsuga menziesii y Pinus ponderosa (los tres muy diversificados en un conjunto de variedades merecedoras de rango taxonómico superior y con un comportamiento ecológico y sucesional muy diferente) se extienden en una buena parte de la superficie potencial ocupada por estos bosques, que representa 7% de la cubierta vegetal de Canadá y los Estados Unidos. Otras herbáceas, en particular algunas saprofitas y hemiparásitas (Sarcodes sanguinea, Pyrola sp., Orthilia sp., Chimaphila sp.) pueden ser también un enlace entre los diferentes bosques, los cuales, por otra parte, comparten una historia común a través del Terciario (Axelrod 1976; Axelrod y Raven 1985). La primera denominación y clasificación científica de estos bosques se debe a Merriam (1890), quien los incluyó en sus biozonas Canadiense, Canadiense de Transición y Hudsoniana, dentro de las cuales distinguió diferentes tipos de bosques cuyas denominaciones, más o menos modificadas, siguen siendo utilizadas actualmente: «Montane Forests of Alta and Baja California» (Barbour y Christensen 1993), «Mixed Conifer Forest» (Barbour 1988; Rundel et al. 1988 Thorne 1988) y «Sierran Montane Conifer Forest» (Pase 1982). Extendidos desde el sur de las Cascadas, por el norte, hasta la Sierra de San Pedro Mártir, cuyos bosques de coníferas constituyen el límite meridional de la clase, resulta difícil encontrar un árbol característico cuya distribución se ajuste por completo al enorme área ocupada por este tipo de bosques, una superficie que representa tan sólo en California 14 % de la superficie estatal (Barbour 1988). De acuerdo con Peinado et al. (1994b), Pinus ponderosa var. ponderosa cumple esta condición, aunque no alcanza los bosques supramediterráneos martirenses, donde es sustituido por Pinus 96 El bosque de coníferas jeffreyi, que también presenta un área de distribución amplia pero más restringida desde el punto de vista ecológico, principalmente por su preferencias en zonas de más baja precipitación o en las que existe algún tipo de sequía edáfica. Esta clase fitosociológica ha tomando en cuenta a Pinus lambertiana por ser un árbol que presenta un área de distribución prácticamente coincidente con la zona de clima mediterráneo de California y que, aunque fundamentalmente supramediterráneo, alcanza también el oromediterráneo. Por otra parte, Arctostaphylos patula ssp. platyphylla es un taxa característico de la clase y de los llamados «chaparrales montanos» que son comunidades permanentes o etapas de sustitución de los bosques maduros climatófilos. En los casos más complejos de otros bosques de confieras puede existir una estructura en cuatro estratos pero, por lo general, el bosque es abierto y fácilmente transitable. El estrato herbáceo suele ser insignificante salvo en las proximidades de zonas encharcadas. En los bosques maduros los árboles del estrato dominante son coníferas monopódicas, de porte generalmente piramidal (excepto viejos ejemplares de Sequoiadendron giganteum, Juniperus australis), cuya altura media oscila entre los 30 y los 60 m, con diámetros de un metro a la altura del pecho y con copas entrelazadas, lo que representa una cobertura del 60 al 80%. En las posiciones más xéricas o en las zonas de más baja precipitación, como en las montañas martirenses, los árboles son más bajos (12-15 m), y el aspecto del bosque más abierto, fenómeno también observado en San Bernardino (Minnich 1976). Las excepciones aparecen también para algunas coníferas como Pinus lambertiana, que supera con facilidad los 65 m, o Sequoiadendron giganteum, que alcanza los 90 m. Asociados con los árboles dominantes aparecen frecuentemente sus parásitos de ramas y hojas: Arceuthobium californicum sobre Pinus lambertiana, Arceuthobium juniperinum subsp. libocedri sobre Calocedrus decurrens, Phoradendron bolleanum y Arceuthobium abietinum sobre el género Abies. El siguiente estrato (5-10 m de alto) no es continuo, se trata de individuos aislados, salpicados bajo el dosel de coníferas, de forma que su cobertura es prácticamente insignificante (Quercus kelloggii, Quercus chrysolepis). El estrato arbustivo, por el contrario, suele estar mejor desarrollado y puede llegar a contribuir de forma sustancial a la cobertura, aunque difícilmente Descripción de la vegetación 97 supera el 25%; son muy característicos los nanofanerófitos postrados de los géneros Arctostaphylos, Ceanothus, Rhamnus spp, Ribes y Symphoricarpos, evidentemente adaptados a soportar el peso de la nieve invernal. Aunque se mencionan coberturas del 25% para el estrato herbáceo (Vankat y Major 1978), por lo general es mucho más bajo (5-10%), predominando especies perennes, algunas de ellas parásitas o hemiparásitas. Son géneros comunes Clintonia, Galium, Iris, Lupinus, Osmorrhiza, Pyrola, Sarcodes, Orthilia y Viola. Es también muy discutido el papel del fuego en el mantenimiento de la estructura original del bosque que habría sido modificada por la política de supresión de fuegos de los últimos decenios (Barbour 1988); no obstante, tal política no se aplicó en Baja California y la estructura de los bosques no es muy diferente. Como otras coníferas californianas, las especies dominantes en esta clase pertenecen al grupo de árboles resistentes e incluso favorecidos por el fuego, como P. lambertiana, P. ponderosa, P. jeffreyi (McCune 1988), y otras coníferas como Abies o Sequoiadendron (Barbour 1988). Cuando el ombroclima es más bajo, lo que regularmente sucede en las provincias Californiano-Meridional y Martirense, así como en las vertientes orientales del eje Sierra Nevada-Cascadas meridionales, Quercus kelloggii no forma comunidades definidas, sino que ocupa posiciones de umbría, por lo general asociado con Abies concolor. En estas zonas de las provincias Californiano-Meridional y Martirense y las vertientes orientales del eje Sierra Nevada-Cascadas, donde la precipitación es más escasa (ombroclima seco), la dominancia corresponde a pinares de P. jeffreyi, acompañados latitudinalmente de Cercocarpus ledifolius var. intermontanus o de sabinas (Juniperus australis). Por descenso de las precipitaciones, P. jeffreyi es reemplazado por pre-bosques semiáridos de la clase Ephedro viridis-Juniperetea osteospermae. En la provincia Martirense no existe un piso oromediterráneo tan claramente definido como en otras montañas californianas, aunque sí hay comunidades de Pinus contorta var. murrayana (Potentillo rimicolae-Pinetum murrayanae), localizadas en zonas con prolongada inundación y permanencia del agua procedente del deshielo. Tanto este pino como Populus tremuloides muestran claras apetencias por lugares húmedos como vaguadas y depresiones (Barbour y Christensen 1993), lugares que, por otra parte, han sido señalados como zonas donde son muy frecuentes las inver- 98 El bosque de coníferas siones climáticas (Haller 1959, 1961), por lo que en San Pedro Mártir pudiera haber oromediterráneos locales indicados por Pinus contorta var. murrayana. Los bosques de pino, pino-sabina y pino-abeto que constituyen la clímax en el piso supramediterráneo de la región Californiana, se encuentran bajo ombroclima seco a subhúmedo. A diferencia de otras comunidades de la clase, las de este orden suelen estar dominadas fisionómicamente por Pinus jeffreyi y ciertas comunidades edafoxerófilas mesomediterráneas propias de sustratos ultrabásicos Pinus jeffreyi, sin ser característica exclusiva, tiene su óptimo en las asociaciones de este orden. Las diferencias ecológicas de este pino con su taxonómicamente próximo P. ponderosa han sido tratadas por diferentes autores (Barbour 1988, McCune 1988, Barbour y Christensen 1993). Todos están de acuerdo en que Pinus jeffreyi domina cada vez más el bosque de coníferas a medida que descienden las precipitaciones, hasta el punto que en los extremos menos lluviosos del área de la clase, como en la provincia Californiano-Meridional y en la Martirense, donde sustituye completamente a P. ponderosa. Por tanto, P. jeffreyi es más resistente a la sequía que P. ponderosa, como lo demuestra su área de distribución, coincidente con las zonas menos lluviosas. Su aparición en las zonas altas de la vertiente occidental de la Sierra Nevada obedece más al descenso en precipitaciones que tiene lugar una vez superada altitudinalmente la zona de mayor descarga de lluvias que a la supuesta resistencia al frío postulada por Haller (1959), Thorne (1988) y Yeaton (1983). La mayor resistencia a la sequía de este pino es también la razón de que sea dominante en sustratos ultrabásicos (dunitas, serpentinas y peridoditas), donde los pinares actúan como comunidades permanentes edafo-xerófilas en áreas de ombroclima subhúmedo-húmedo. ASOCIACIÓN SYMPHORICARPO OREOPHILI-PINETUM JEFFREYI PEINADO ET AL. (1994B) Características. Symphoricarpos oreophilus, Abies concolor. Estructura. Bosque de coníferas dominado por Pinus jeffreyi (20-30 m de alto), cuyas copas no llegan a juntarse, lo que produce un sotobosque abierto, heliófilo, en el que predominan arbustos rastreros o poco elevados (0,51,5 m) como Arctostaphylos patula, Symphoricarpos oreophilus, Holodiscus Descripción de la vegetación 99 microphyllus var. sericeus, Rhamnus californica, y un número escaso de herbáceas nemorales (Monardella macrantha, Phlox austromontana, Bromus ciliatus, Silene platyota). La mayor densidad y diversidad del bosque se alcanza en las laderas orientadas al norte o en altitudes superiores a los 2,200 m; en estas situaciones penetran Abies concolor (hasta 30 m de alto), Pinus lambertiana (30 m) y Calocedrus decurrens (hasta 45 m). Por lo general, C. decurrens parece preferir suelos con alguna humedad edáfica, denunciando la ecotonía con los bosques sobre suelos temporalmente encharcados (Potentillo rimicolae-Pinetum murrayanae). Ecología. Piso supramediterráneo con ombroclima seco de la alta montaña bajocaliforniana. Fitogeografía. Asociación endémica del sector Martirense SINFITOSOCIOLOGÍA. ETAPA MADURA DE LA SERIE SUPRAMEDITERRÁNEA SECA, MARTIRENSE DEL PINO DE JEFFREY Observaciones. El piso supramediterráneo de las altas montañas del oeste de Norteamérica está dominado por bosques de coníferas (Pinus ponderosa, P. jeffreyi, P. lambertiana, Abies concolor, Calocedrus decurrens, etc.) que, sobre todo en el horizonte inferior del piso, se mezclan con caducifolios (en particular con Quercus kelloggii). P. ponderosa y A. concolor son los elementos comunes desde las Rocosas a las montañas del sur de California; en tanto que P. jeffreyi y A. concolor son elementos comunes en la Sierra de San Pedro Mártir. Estos pinares en las laderas más secas están en contacto con bosques mesomediterráneos semiáridos de P. monophylla. La escasez de precipitaciones determina la inexistencia de caducifolios en la provincia Martirense, (por ejemplo Quercus kelloggii), y la presencia en el piso supramediterráneo de Quercus chrysolepis, un elemento típicamente mesomediterráneo en otras provincias de la región Californiana donde la precipitación alcanza valores del subhúmedo. ASOCIACIÓN POTENTILLO RIMICOLAE-PINETUM MURRAYANAE PEINADO ET AL. 1994B. Características. Potentilla wheeleri var. rimicola, Pinus contorta var. murrayana, Populus tremuloides. 100 El bosque de coníferas Estructura, ecología y fitogeografía. Comunidad abierta dominada por P. contorta var. murrayana y Pinus jeffreyi, cuyo sotobosque carece de un estrato arbustivo bien desarrollado, en cambio existe un estrato herbáceo perenne (Carex brevipes, Achillea millefolium var. californica, Potentilla rimicola) que confiere a la asociación el aspecto de un parque, muy característico en las zonas de Vallecitos y La Grulla en la Sierra de San Pedro Mártir. Potentillo rimicolae-Pinetum murrayanae es una asociación edafohigrófila que vive sobre protofluvisoles originados por la acumulación de agua del deshielo en microdepresiones (Vallecitos) o por la existencia de cursos poco importante de agua; en estos últimos Populus tremuloides es un componente habitual de la asociación. Por lo estricto de su hábitat la asociación está muy restringida en el piso supramediterráneo superior de la Sierra de San Pedro Mártir, donde aparece en áreas puntuales dentro de la potencialidad del pinar Symphoricarpo oreophili-Pinetum jefreyii. También son comunes en toda la región Californiana (meadow forests) los bosques de ecología similar dominados por Pinus murrayana sobre un césped de higrófilas. Ésta una asociación endémica del sector Martirense, bastante desviante de otras descritas para el orden, pero que probablemente puedan unirse sintaxonómicamente una vez que se hayan descrito sus vicariantes septentrionales. Descripción de la vegetación 101 102 El bosque de coníferas Glosario Abiótico: desprovisto de vida, no vivo. Antropogénico: ocasionado o producido por la acción humana; antropógeno. Área: en biogeografía (corología), conjunto de lugares en los que se hallan individuos de la misma especie o comunidades. Se distingue entre áreas continuas o discontinuas (disyuntas) para expresar su modo de distribución. Área protegida: aquélla que haya sido designada o regulada y administrada a fin de alcanzar objetivos específicos de conservación. Árido: clima o hábitat con precipitación pluvial baja, menor a 250 mm anuales, evaporación que excede a la precipitación y vegetación escasa. Asociación: unidad fundamental y básica de las comunidades vegetales, que posee unas peculiares cualidades florísticas (especies características y diferenciales), ecológicas, fitogeográficas, dinámicas e históricas. Bioclimatología: ciencia ecológica que trata de clarificar las relaciones existentes entre organismos vivos y clima. Bioindicador: se dice tanto de especies como comunidades que pueden ser utilizados para poner de relieve propiedades del medio o unidad de lugar. Por extensión puede hablarse de bioindicadores geográficos, climáticos, edáficos, etc. Se utiliza también como sustantivo. Biológico: referente a los organismos vivos o a los procesos de vida. Biótico: relativo a la vida y a los organismos vivos; causado o producido por organismos vivientes o comprendido por estos. Biotopo: unidad más pequeña de la biosfera o de un hábitat que puede ser delimitado mediante fronteras convenientes. Bosque: área relativamente grande de árboles que crecen con gran proximidad entre ellos. Glosario 103 Cenozoico: era geológica (aproximadamente. 65-0 millones de años antes del presente) que comprende los periodos Cuaternario y Terciario; también conocida como la Edad de los mamíferos. Clima: conjunto de condiciones atmosféricas propias de un área o territorio. Clima Mediterráneo: característica climática propia de la Cuenca Mediterránea, y otras partes del mundo, con un período de lluvias (invierno-primavera) y otro de sequía (verano). Clímax: etapa final de equilibrio en la sucesión de una comunidad (e.g. vegetal) que representa territorialmente la etapa de máximo biológico estable. Se puede emplear también como expresión del ecosistema maduro, y como la etapa final o asociación estable y madura de una serie. Su adjetivo es climácico. Cliserie: la serie de asociaciones vegetales determinadas por la sucesión de climas en sentido altitudinal o latitudinal. Comunidad vegetal: conjuntos más o menos homogéneos de plantas, pertenecientes a distintos táxones que ocupan un área y medio determinados. Corología: ciencia que estudia las causas de la distribución y localización de las especies y ecosistemas sobre la Tierra. Cripto: prefijo que alude a algo que está oculto o poco manifiesto; ejemplo criptoprecipitaciones («precipitaciones ocultas»). Chaparral: nombre común a un tipo de vegetación característica de la región Mediterránea, Californiana y Bajacaliforniana, representada por una flora arbustiva, siempreverde y pirófila. Deciduo: plantas que tiran sus hojas. Desinencia: letra o letras que subsiguen al radical de los vocablos. En botánica y de acuerdo a las Reglas Internacionales de Nomenclatura: divisiones, -phyta; clases, opsida; ordenes, -ales; familias, -aceae. En fitosociología se emplea de acuerdo al Código de Nomenclatura Fitosociológica: clases, –etea; ordenes, -etalia; alianzas, ion; asociaciones, -etum. Ecosistema: sistema complejo formado por una trama de elementos físicos (biótopo) y biológicos (comunidades de organismos o biocenosis). Ecosistema vegetal: sistema biológico que integra los factores del medio y las comunidades vegetales. Endémico: especies de flora o de fauna exclusivas de un área geográfica particular. Esclerófilo: plantas de hojas duras, coriáceas (dureza como el cuero). Escurrimiento: parte de la precipitación caída en una que no se infiltra y desagua superficialmente. 104 El bosque de coníferas Estación: la suma de factores ecológicos que constituyen el medio de una comunidad. Véase medio. Eoceno: época geológica dentro del periodo Terciario (hace aproximadamente. 5438 millones de años). Erosión: pérdida progresiva del suelo que se produce en los terrenos por acción físico-química del agua, viento y agentes biológicos. Facies: término empleado en geobotánica con diferencia de significados. Lorenz lá introdujo en 1863 aplicándolá a las “formaciones o subdivisiones de ellas consideradas desde el punto de vista de la localización y de los factores físicos que las condicionan”. Fisiográfico: perteneciente a las características geográficas de la superficie terrestre; fisiografía. Fitogeografía: ciencia que estudia la distribución y origen geográfico de las plantas. Sinónimo de geografía botánica. Flora: conjunto de plantas de un país o territorio cualquiera. Obra que trata de ellas, las enumera, las describe, e indica donde se desarrollan. Cuando no se describen las plantas, en lugar de flora es más correcto emplear otro término: catálogo, enumeración, lista, etc. Freático: agua no superficial situada a cierta profundidad del suelo. Freatófita: planta que depende del agua freática y no de la humedad del suelo (e.g. sauces, álamos, mezquites). Geología: ciencia que estudia la estructura, procesos y cronología de la Tierra. Hábitat: lugar o tipo de ambiente en el que existen naturalmente un organismo o una población. Inframediterráneo: piso bioclimático por la abundancia de elementos endémicos y sensibles a las heladas, algunos de claro origen neotropical (Agaváceas, Cactáceas, etc). Tradicionalmente ha sido considerado como un área transicional entre los ecosistemas mediterráneos y desérticos. Jerarquía: característica de un sistema general integrado que comprende dos o más niveles; el más alto controla, en cierta medida, las actividades de los niveles inferiores. Matorral costero suculento: tipo de vegetación costera californiana que se caracteriza por presentar una flora subarbustiva y aromática; son caducifolias de verano. Se distribuye principalmente en el noroeste de Baja California. Medio: suma de factores que integran una unidad de lugar. Hay que distinguir entre el medio geográfico, referido a sus características, y medio estacional, como la Glosario 105 suma de los factores ecológicos naturales que inciden y condicionan dicho lugar. El medio antropógenico sería el profundamente modificado por el hombre y sus actividades. Véase estación. Mioceno: época geológica situada en le periodo Terciario (hace aproximadamente. 26-25 millones de años). Ombroclima: la parte del clima que se refiere a las lluvias o precipitaciones. La cantidad de lluvia que cae en una localidad se expresa en litros por metro cuadrado o en milímetros de altura, que es el mismo número. En la región mediterránea se distinguen seis tipos de ombroclima según sea la media anual en mm: árido (inferior a 200), semiárido (200-350), seco (350-600), subhúmedo (600-1,000), húmedo (1,000-1,600), e hiperhúmedo (superior a 1,600). Paisaje: parte de la superficie terrestre que en su imagen externa y en la acción conjunta de los fenómenos que lo constituyen presenta caracteres homogéneos y cierta unidad espacial básica. Pastizal: área de vegetación dominada por pastos herbáceos («zacate»); algunas veces se denomina así a cualquier tipo de vegetación donde domina la hierba. Pirófila: vegetación o flora adaptada al fuego (por ejemplo, chaparral). Pisos bioclimáticos: cada uno de los tipos o grupos de medios que se suceden en un gradiente altitudinal o latitudinal. En la práctica se delimitan en función de los factores climáticos y por el cambio en las comunidades vegetales cambiantes. Pisos de vegetación: cada uno de los complejos o series de vegetación que se escalonan en un gradiente altitudinal. Planta perenne: dicese de la planta que vive tres años o más. Pleistoceno: época geológica del periodo Cuaternario (aproximadamente. 1.6-0.001 millones de años antes del presente); también denominada Gran Edad del hielo. Provincia: unidad tipológica de la biogeografía (corología) intermedia entre la región y el sector. Es un territorio extenso que posee muchas especies propias e incluso paleoendemismos. Recursos biológicos: los recursos genéticos, los organismos o partes de ellos, las poblaciones, o cualquier otro tipo de componente biótico de los ecosistemas de valor o utilidad real o potencial para la humanidad. Región: unidad biogeográfica de rango superior a provincia e inferior a reino floral. Es un territorio muy extenso que posee especies, géneros e incluso familias propias. Regionalización: división de un territorio. Ripario: que vive o está situado sobre los bancos de ríos y las corrientes. 106 El bosque de coníferas Sector: unidad tipológica de la biogeografía situada entre la provincia y el distrito. Suele ser un territorio bastante extenso que posee algunas especies (táxones) y asociaciones propias. Siempreverde: plantas con sus hojas verdes todo el año. Sucesión: proceso natural por el que se sustituyen unas comunidades vegetales o estadios a otras dentro de la misma unidad del lugar. Suculento: plantas con tejido especializado carnoso en la raíz o el tallo que almacena agua. Taxón: en la sistemática de las plantas o taxonomía vegetal ,cualquiera de los rangos o tipos que se reconocen. La unidad básica de esta tipología es la especie. Taxonomía: ciencia que describe, nombra y clasifica organismos (plantas y animales). Termoclima: la parte del clima que se refiere a las temperaturas; en la región mediterránea se puede hablar de cinco tipos de termoclima, según sea la media anual en grados centígrados: cálido (superior a 16), templado (de 16 a 12), frío (de 12 a 8), muy frío (de 8 a 4) y extremadamente frío (inferior a 4). Termomediterráneo: piso bioclimático con termicidad propia de chaparrales esclerófilos. Textura: el carácter o estructura de un sedimento o de una superficie. Topoforma: forma, variedad o subespecie geográfica. Se aplica también desde el punto de vista topográfico. Topografía: referente a todas las características de la superficie de un área geográfica, tanto las naturales como las restantes de la actividad humana. Vegetación: conjunto de plantas que pueblan un área determinada y que ejercen entre sí múltiples influencias. Vegetación actual: ver vegetación real. Vegetación clímax: vegetación relativamente estable en equilibrio con su ambiente y con una buena reproducción de las plantas dominantes. Vegetación potencial: comunidad vegetal estable que existiría en un área dada como consecuencia de la sucesión geobotánica si el hombre dejase de influir y alterar los ecosistemas vegetales. Vegetación real: comunidad vegetal que existe en un lugar dado sometida a la influencia del medio estacional y antropógena. Es sinónimo de vegetación actual. Vertiente: planos en declive que divergen de las crestas o de los interfluviales y que limitan un valle. Xerófilo: calificativo en un sentido general que se aplica a las plantas que viven en ambientes secos. Glosario 107 108 El bosque de coníferas Bibliografía Alcaraz, F. 1991-1995. Programa BIOCLIMA. Universidad de Murcia, España. ——— 1999. Manual de teoría y práctica de geobotánica. ICE-Universidad de Murcia, España. Álvarez, M. 1983. Climatología de la Sierra de San Pedro Mártir. Presentado en el 15o. Simposio Anual del Desert Fishes Council, Death Valley, California. Noviembre de 1983. Arno, F.S. Discovering Sierra trees. Yosemite Association and Sequoia Natural History Association, in cooperation with the National Park Service, USDI. Axelrod, I. D. 1975. Evolution and biogeography of Madrean-Tethyan sclerophyll vegetation. Annals Botanical Garden 62: 280-334. ——— 1976. History of the Coniferous Forests, California and Nevada. University of California Press, Berkeley. Pp. 1-62. ——— 1988. Outline history of California vegetation. En: M.G. Barbour y J. Major (eds.). Terrestrial vegetation of California. California Native Plant Scociety, Davis. Pp. 139-194. Axelrod, I.D. y P.H.Raven. 1985. Origins of the cordilleran flora. Journal of Biogeography 12: 21-47. Azuela, A., J. Carabias, E. Provencio y G. Quadri (coords.).1993. Desarrollo sustentable: hacia una política ambiental. Universidad Nacional Autónoma de México. México, D.F. Barbour, G.M. 1988. California uplands forest and woodlands. En: M.G. Barbour y W.D. Billings (eds.). North American Terrestrial Vegetation. Cambridge University Press, New York. Pp. 131-164. Barbour, G.M y W.D. Billings.1988. North American terrestrial vegetation. Cambridge University Press, Cambridge. Bibliografía bibliog.pmd 109 13/07/2005, 12:35 109 Barbour, G.M. y N.L. Christensen. 1993. Vegetation. En: Flora of North America Editorial Commitee (eds.). Flora of North America, north of Mexico. Vol. 1. Oxford University Press, New York-Oxford. Pp. 97-131. Barkman, J.J., J. Moravec y S. Rauschert. 1988. Código de Nomenclatura Fitosociológica (traducido por J. Izco y M.J. del Arco-Aguilar). Opuscula Botanica Pharmaciae Complutense 4: 5-74. Barry, W.J. 1991. The potential for a transborder Biosphere Reserve representing the peninsular range ecological region of Baja and Alta California. Commision of the Californias, 46 th General Assembly. Cabo San Lucas, Baja California Sur, de abril de12, 1991. Beauchamp, R.M. 1986. A flora of San Diego County, California. Swettwater River Press, National City, California. Brown, E.D. 1982. Great basin conifer woodland En: David E.Brown (ed.), Biotic communities of american southwest, United States and Mexico. Desert Plant 4 (1-4): 52-57. Clements, F.E. 1916. Plant sussession: an analysis of the development of vegetation. Carnegie Inst. Wash. Pub., Washington,D.C. CONABIO. 1998. Regiones prioritarias para la conservación en México. Comisión nacional para el Uso y Conocimiento de la Biodiversidad. México, D.F. Christensen, N.L. y C.A. Muller. 1975. Relative importance of factors controolling germination and seedling survival in Adenostoma chaparral. The American Midland Naturalist 93:71-78. Critchfield,W.y E.Little,Jr.1966.Geographic distribution of the pines of the world.Miscellaneous Publication 991. Forest Service, Departament of Agriculture. Washington, DC. Cruz, V. D. 2001. Diagnosis de las formas biológicas y de vida de la flora vascular de la Sierra San Pedro Mártir, Baja California. Tesis de Licenciatura, Facultad de Ciencias, Universidad Autónoma de Baja California. Ensenada, B.C. Dansereau, p. 1957. Biogeography: an ecological perspective. The Ronald Press Company, New York. Daubenmire, F.R. 1979. Ecología vegetal: tratado de autoecología de plantas. Limusa, Mexico. Delgadillo, J. 1987. Aspectos florísticos y ecológicos de las Sierras Juárez y San Pedro Mártir, Baja California; Memorias del X Congreso Mexicano de Botánica, Guadalajara, Jal., 27 septiembre a 3 octubre 1987. ——— 1995. Introducción al conocimiento bioclimático, fitogeográfico y fitosociológico del suroeste de Norteamérica y México. Tesis Doctoral, Universidad de Alcalá de Henares, España. 110 El bosque de coníferas bibliog.pmd 110 13/07/2005, 12:35 ——— 1998. Florísitica y ecología del norte de Baja California. Universidad Autónoma de Baja California. Mexicali, B.C., México. ——— 1999. Vegetación y análisis fitogeográfico de la flora vascular de la Sierra San Pedro Mártir, Baja California. Reporte final del Proyecto L077. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Agosto, 1999. Duffield, W.F. y W.C. Cumming.1974. Does Pinus ponderosa occur in Baja California? Madroño 10: 22-24. Dunn, L.E., F. M. Shropshire, L.C. Song y H.A. Mooney. 1976. Water factor an convergent evolution in Mediterranean type vegetation. En: O.L. Lange, L.Happen, E. D. Schulze (eds.). Water and plant life problems and modern approaches. Springer Verlag, New York. Eguiluz, P.T. 1985. Origen y evolución del género Pinus (con referencia especial a los pinos mexicanos). Dasonomía Mexicana 3(6): 5-31. Escoto, R. M. 1994. Balance del agua del suelo en tres sitios de estudio en la Sierra de San Pedro Mártir, Baja California. Tesis de Licenciatura, Facultad de Ciencias, Universidad Autónoma de Baja California, Ensenada, B.C. Estrada, A. 1995. Los Kiliwas y la Sierra de San Pedro Mártir. En: E. Franco-Vizcaíno y H. de la Cueva (eds.). Estrategias de Conservación para la Sierra de San Pedro Mártir. CICESE-UCMEXUS, Ensenada, B.C., 4-5 de diciembre de 1995. Flores, M., G. Jimenez, J. Madrigal, X. Moncayo y F. Takaki. 1971. Memoria del mapa de tipos de vegetación de la República Mexicana. Secretaría de Recursos Hidráulicos. México, D.F. Foster, W.J. 1991. La misión de San Pedro Mártir de Verona: una breve historia y evaluación de potencial para el turismo interpretativo dentro de una Reserva de la Biosfera. En: E. Franco-Vizcaíno y J. Sosa-Rámirez (eds), Memorias de la conferencia internacional sobre el potencial de la cordillera peninsular de las Californias como Reserva de la Biosfera. CICESE-UABC-UC-CSU. Ensenada, B.C., 18y 19 de marzo de 1991. Font-Quer, P. 1979. Diccionario de botánica. Labor. Barcelona, España. García, E. 1981. Modificaciones al sistema de clasificación climática de Köppen (para adaptarlo a las condiciones de la República Mexicana). Tercera edición. UNAM, México. García, E. y P.A. Mociño. 1968. Los climas de Baja California. Memoria 1966-1967. México, D.F. García, M.T. y M.A. Barrios. 2001. Glosario de términos biológicos. Instituto Politécnico Nacional. México, D.F. 97 pp. Bibliografía bibliog.pmd 111 13/07/2005, 12:35 111 Gastil, L.G., R.P. Philips y E.C. Allison. 1975. Reconnoissance geology of the State of Baja California. The Geological Society of America, Inc., Colorado. Gastil, L.G. 1991. Geología de la Sierra de San Pedro Mártir. En: E. Franco-Vizcaíno y J. Sosa-Rámirez (eds.). Memorias de la conferencia internacional sobre el potencial de la cordillera peninsular de las Californias como Reserva de la Biosfera. CICESEUABC-UC-CSU. Ensenada, B.C., 18-19 de marzo 1991. Gobierno del estado de Baja California. 1996. Convenio de Concertación para constituir el Comité Técnico del Parque Nacional Sierra de San Pedro Mártir. SFA, SEMARNAP. Mimeo. Gouldman, A. E. 1916. Plant record of an expedition to Lower California. United States National Herbarium. Contribution Smithsonian Institute. Vol. 16, part. 14. Gentry, S.H. 1978. The agaves of Baja California. Occasional Paper California Academy of Sciences, No. 130. San Francisco, California. Gutierrez, R.J., S. Camacho y R. Naranjo. 1983. Glosario de recursos naturales: agua, suelo y vegetación. México, D.F. Haller, J.R. 1959. Factors affecting the distribution of ponderosa and jeffrey pines in California. Madroño 15 : 65-96. ——— 1961. Some recent observations on ponderosa, jeffrey and washoe pines in northeastern California. Madroño 16: 126-133. Hanes, L.T. 1976. Vegetation types of the San Gabriel Mountains. En: L. J. Davis (ed.). Plant communities of Southern California, California Native Plant Society. Pp. 6576. Hasting, J.R. y R.M. Turner. 1965. Seasonal precipitation regimes in Baja California, México. Geografiska Annals 47: 204-223. Hernandez, M.A., A. Mancera, F. Martínez, R. Sainas y D. Zavala. 1977. Observaciones ecológicas, fitosanitarias (plagas y enfermedades) y sobre aprovechamientos industriales en las Sierras de Juárez y San Pedro Mártir, en el estado de Baja California Norte. Ciencia Forestal 2(9): 3-38. Hickman, C. J. (ed.). 1993. The Jepson Manual: higher plants of California. University of California Press, Berkeley. INEGI. 2001. Síntesis de información geográfica del Estado de Baja California. Institu- to Nacional de Estadística, Geografía e Informática. Aguascalientes, México. Iturbe, C. J., C. M. Olivas y R. I. Castillo. 1984. Uso multiple del bosque en el Parque Nacional Sierra de San Pedro Mártir, Baja California. Tesis Profesional de Licenciaura Ingeniero Agrónomo. Universidad Autónoma de Chapingo. Chapingo, México. 112 El bosque de coníferas bibliog.pmd 112 13/07/2005, 12:35 Jaroslav, M. 1992. Is the Zürich-Montpellier approach still unknown in vegetation science of the English-speaking countries? Journal of Vegetation Science 3: 277278. Keeley, J y S.C. Keeley. 1988. Allelopathy and the fire-induced herb cycle. En: S.C. Keeley (ed.). The California chaparral: paradigms reexamined. Natural History Museum of Los Angeles County. Science Series 34: 65-72. Keeley, J. y P. Zedler. 1998. Evolution of life histories in Pinus. En: M.D. Richardson (ed.). Ecology and biogeography of pines. Cambridge University Press, Cambridge. 249 pp. Kovalchik, B.L. y L.A. Chitwood. 1990. Use of geomorphology in the classification of riparian plant associations in mountainous landscapes of Central Oregon. Forest Ecology and Management 33/34: 405-418. Laacke, R. 2004. Abies concolor; white fir. www.na.fs.fed.us/spfo/pubs/silvics_manual/ Volume_1/abies/concolor.htm. Leopold, A.S. 1950. Vegetation zones of Mexico. Ecology 31: 507-518. Lincoln, J.R., G.A. Boxshall y P.F. Clark. 1996. Diccionario de ecología, evolución y taxonomía. Fondo de Cultura Económica, México, D.F.488 pp. Long, G. 1990. Sur la distribution de la vegetation mediterranenne: de la cote du Pacifique aux cortes de la Sierra de San Pedro Mártir, Basse Californie, Mexique. Ecologia Mediterránea 16 : 269-277. Major, J. y D.W. Taylor. 1988. Alpine. En: Terrestrial vegetation of California. California Native Plant Society. Special Pub. 9. Martínez, M. 1945. Las pináceas mexicanas. Anales del Instituto de Biología 16: 1-345. McCune, B. 1988. Ecological diversity in North American pines. American Journal of Botany 75 (3): 353-368. Meling, D. 1991. La ganadería en la Sierra de San Pedro Mártir. En: E. Franco-Vizcaíno y J. Sosa-Rámirez (eds). Memorias de la conferencia internacional sobre el potencial de la cordillera peninsular de las Californias como Reserva de la Biosfera. CICESE-UABC-UC-CSU. Ensenada, B.C., 18-19 de marzo de 1991. Meling, E. 1995. La ganadería en la Sierra de San Pedro Mártir. En: E. Franco-Vizcaíno y H. de la Cueva (eds.). Estrategias de Conservación para la Sierra de San Pedro Mártir. CICESE-UCMEXUS, Ensenada, B.C., 4-5 de diciembre de 1995. Merrian, C.H. 1890. Life-zones and crop-zones of the United States. US Departament of Agricultura. Division of Biological Survey Bulletin 10:1-79. Minnich, R.A. 1976. Vegetation of the San Bernardino Mountains. En: J.D. Latting (ed.). Plant communities of Southern California. California Native Plant Society. Pp. 99-124. Bibliografía bibliog.pmd 113 13/07/2005, 12:35 113 ——— 1982. Pseudotsuga macrocarpa in Baja California? Madroño 2(1): 22-31 ——— 1983. Fire mosaics in southern California and northern Baja California. Science 219: 1287-1294. ——— 1985. Evolutionary convergense or phenotypic plasticity?, responses to summer rain by California chaparral. Physical Geography 6 (3): 272-287. ——— 1986. Range extensions and corrections for Pinus jeffreyi and P. coulteri (Pinaceae) in northern Baja California. Madroño 33(2): 144-46. ——— 1987. The distribution of forest trees in northern Baja California, Mexico. Madroño 34(2): 272-287. ——— 1995. Propuesta de Reserva de la Biósfera para la Sierra de San Pedro Mártir.En: E. Franco-Vizcaíno y H. de la Cueva (eds.). Estrategias de Conservación para la Sierra de San Pedro Mártir. CICESE-UCMEXUS, Ensenada, B.C., 4-5 diciembre de 1995. Minnich, R.A., E. Franco-Vizcaíno, J. Sosa-Ramírez, J.H. Burk, W. J. Barry, M. G. Barbour y H. de la Cueva. 1994. El potencial de la Sierra de San Pedro Mártir como Reserva de la Biósfera. Reporte al Proyecto Áreas Naturales Protegidas de México. 51 pp. Minnich, R.A., E. Franco-Vizcaíno, J. Sosa-Ramírez, J.H. Burk, W. J. Barry, M. G. Barbour y H. de la Cueva. 1997. A land above: protecting Baja California´s Sierra San Pedro Mártir within a Biosphere Reserve. Journal of the Southwest 39(4): 611-695. Minnich, R.A. y E. Franco-Vizcaíno. 1997a. La vegetación mediterránea del norte de Baja California. Fremontia 25(3): 3-12. ——— 1997b. La protección de la vegetación y los regímenes de incendios de la Sierra de San Pedro Mártir en Baja California. Fremontia 25(3): 13-21. ——— 1998. Land of chamise and pines: historical account ans current status of vegetation in northern Baja California. University of California, Publications in Botany 80: 1-166. Minnich, R.A., M. G. Barbour, J.H. Burk y J. Sosa-Ramírez. 2000. Mixed-conifer forest under unmanaged fire in the Sierra San Pedro Mártir, Baja California, México. Journal of Biogeography 27: 105-130. Miranda, R.F., M. Alvarez y J.C. Leal. 1997. Precipitación pluvial en Baja California en los alrededores de San Pedro Mártir. Mimeo. Mooney, A.H. y A.T. Harrison. 1972. The vegetational gradiente on the lower slopes of Sierra San Pedro Mártir, in northwestern Baja California. Madroño 21: 439-445. Moran, R. 1977. Botany of Sierra San Pedro Martir trip. San Diego Natural History Museum, Mimeo. Oberbauer, T. 1996. Sierra San Pedro Mártir. Fremontia 24(4): 20-24. 114 El bosque de coníferas bibliog.pmd 114 13/07/2005, 12:35 O’Connor, J.E. y Chase. 1989. Uplift of the Sierra San Pedro Mártir Baja California, México. Tectonics 8: 833-844. Pase, P. CH. 1982. Rocky Mountain (Petran) and Madrean Montane Conifer Forest. En: David E. Brown (ed.), Biotic communities of the american southwest-United States and Mexico. Desert Plant 4(1-4). Pase, P. CH y E. Brown. 1982. California coastal scrub and Californian (coastal) chaparral. En: David E. Brown (ed.). Biotic communities of the american southwest-United States and Mexico. Desert plant 4(1-4): 91-99. Passini, F. M., J. Delgadillo y M. Salazar. 1989. L’ecosysteme forestier de Basse Californie: composition floristique: variables ecologiques principales dynamique. Oecologia Plantarum 10(3): 275-293. Peinado, M. y J. Delgadillo. 1990. Introducción al conocimiento fitogeográfico de Baja California, México. Stvdia Botánica 9: 25-39. Peinado, M., F. Alcaraz, J. Delgadillo e I. Aguado. 1994a. Fitogeografía de la península de Baja California, México. Anales del Jardín Botánico de Madrid 51(2): 255-277. Peinado, M., C. Bartolomé, J. Delgadillo e I. Aguado. 1994b. Pisos de Vegetación de la Sierra de San Pedro Mártir, Baja California, México. Acta Botánica Mexicana 29: 1-20. Peinado, M., F. Alcaraz, J.L. Aguirre y J. Alvarez. 1995. Vegetation formations and associations of the zonobiomes along the North American Pacific Coast. Vegetatio 114: 123-135. Peinado, M., J.L. Aguirre y J. Delgadillo. 1997. Phytosociological, bioclimatic and biogeographical classification of woody climax communities of western North America. Journal of Vegetation Science 8: 505-528. Perry, J. Jr. 1991. The pines of Mexico and Central America. Timber Press. Portland Oregon. Raven, H.P. 1973. The evolution of Mediterranean floras: En: F. di Castri y H.A. Mooney (eds). Mediterranean type vegetation: origen and structure. Springer-Verlag, New York. ——— 1988. The California flora: En: M.G. Barbour y J. Major (eds.). Terrestrial vegetation of California. California Native Plant Society, Spec. Pub. 9: 109-138. Raven, H.P. y D.I. Axelrod. 1978. Origen and relationships of the California flora. University of California, Publication in Botany 72: 1-34. Reichenbacher, W.F. 1984. Ecology and evolution of southwestern riparian plant communities. Desert Plants 6(1): 15-22. Reyes, S. 1993. Lluvias en Baja California. Periódico Z, No. 1030: 22A-24A (22 al 29 diciembre 1993). Tijuana, B.C. Bibliografía bibliog.pmd 115 13/07/2005, 12:35 115 Rivas-Martinez, S. 1987. Nociones sobre fitosociología, biogeografía y bioclimatología.En: M. Peinado y S. Rivas-Martínez (eds.). La vegetación de España: Serv. Publ., Universidad de Alcalá de Henares, Madrid. Pp.17-46. Robichaux, R. 1977. Geological history of riparian forest of California. En: A. Sands (ed.). Riparian forest in California: their ecology and conservation. A Symposium Sponsored by Institute of Ecology, University of California, Davis and Davis Audubon Soc., May 14, 1977. Pub. No.15. Rundel, W.P., D.J. Parson y D.T. Gordon. 1988. Montane and subalpine vegetation of the Sierra Nevada and Cascade Ranges.En: M.G. Barbour y J.Major (eds.). Terrestrial vegetation of California. California Native Plant Society. Pp. 559-600. Rundel, W. P. y J. L. Vankat. 1989. Chaparral communities and ecosystems.En: S.C. Keeley (ed.). The California chaparral: paradigms reexamined. Natural History Museum of Los Angeles County. Science Series 34: 127-139. Rzedowski, J. 1978, 1986. La vegetación de México. Limusa, México. ——— 1985. Análisis de la distribución geográfica de las especies del género Populus (Salicaceae). C.R. Soc. Biogéogr. 60(4): 141-150. ——— 1992. Diversidad y orígenes de la Flora Fanerogámica de México. En: Acta Zoológica Mexicana, Vol. especial. La diversidad biológica de Iberoamérica. I. Halffter, G. (comp.) Xalapa, Ver. Pp. 337-359 ——— 1994. Geographical affinities of the riparian trees of Mexico. Mem. Soc. Biogéography IV: 37-44. Salas, L. 1997. El observatorio Astronómico SPM a 30 años de su aniversario. Mesa Redonda “Parque Nacional Sierra de San Pedro Mártir a 50 años”. Parque Nacional Sierra de San Pedro Mártir y Facultad de Ciencias, UABC. 24 abril de1997. Sawyer, J.O., D.A. Thornburgh y J.R. Griffin. 1988. Mixed evergreen forest. En: M.G. Barbour y J. Major (eds.). Terrestrial vegetation of California. Whiley, New York. 359-381 pp. Secretaría de Agricultura y Recursos Hidraulicos. 1977. Inventario Forestal del Estado de Baja California. Resumen de la Publicación No.3 (1968). Dirección General de Inventarios Forestales, SARH, México. Publicación No.43. 24 pp. ——— 1991. Datos climáticos en el Estado de Baja California. División Hidrométrica, Ensenada, Baja California, México. Secretaría de Medio Ambiente y Recursos Naturales. 2000. Áreas naturales protegidas de México con decretos federales. (1899-2000). Instituto Nacional de Ecología, Semarnap, México, D.F. pp. 36-37. ——— 2001. Norma Oficial Mexicana NOM-059-ECOL-2001. 116 El bosque de coníferas bibliog.pmd 116 13/07/2005, 12:35 Sierra, P.A. 1995. Estudio técnico justificativo para el saneamiento forestal: predio Ejido El Bramadero, Municipio de Ensenada, Baja California. 65 pp (+ Anexo B). Swanson, F.J., S.V. Gregory, J.R. Sedell y A.G. Cambell. 1982. Land-water interactions: the riparian zone:En:R.L.Edmonds (ed.).Analysis of coniferous forest ecosystems in the western United States. US/IBP Sunthesis Series, Hutchinson Ross Pub. Co. pp. 267-291. Thorne, F.R. 1985. Further botanical adventures in northwestern Baja California. Crossosoma 11(6): 8-12. ——— 1988. Montane and subalpine forest of the Transverse and Peninsular Ranges. En: M.G. Barbour y J. Major (eds.). Terrestrial vegetation of California. California Native Plant Society. Publ. Spec. 9. Thorne, F.T. y R.V. Moran. 1986. An annotated checklist of vascular plants of the conifer forest of Sierra San Pedro Martir, Baja California, Mexico. Mimeo. Van Devender, T. 1990. Late Quaternary vegetation and climate of the Chiuahuan desert, United States and Mexico. En: J. Betancourt, T. Van Devender y P. Martir. Packrat middens: the last 40,000 years of biotic change. The University of Arizona Press, Tucson, AZ. 104-132 pp. Vale, R.T. 1979. Pinus coulteri and wildfire on Mount Diablo, California, Madroño 26(3): 135-140. Vankat, J.L. y J. Major. 1978. Vegetation changes in Sequoia National Park, California. Journal of Biogeography 5:377-402. Vasek, F.C. y R.F. Thorne. 1988. Transmontane coniferous vegetation. En: M.G. Barbour y J. Major (eds.). Terrestrial Vegetation of California. California Native Plant Society. 797-834 pp. Villaseñor, J.L. y T.S. Elias. 1989. Endemics and conservation in Baja California, Mexico. 1er. Simposio sobre Recursos Vegetales, Escuela Superior de Ciencias Biológicas, Universidad Autónoma de Baja California y Rancho Santa Ana Botanic Garden. Ensenada, Baja California, México. 17-18 de noviembre de 1989. Walter, H. 1985. Vegetation of the Earth and ecological systems of the geo-biosphere. Tercera edición Springer-Verlag Berlin Heidelberg, Berlin. Wells, P.V. 1983. Paleobiogeography of montane islands in the Great Basin since the last glaciopluvial. Ecological Monographs 53(4): 341-382. Warren, G.R., J. Greg y J. Major. 1977. A survey of riparian forest flora and conservation. En: A. Sands (ed.). Riparian forest in California: their ecology and conservation. A Symposium Sponsored by Institute of Ecology, University of california, Davis and Davis Audubon Soc., de 14 mayo de 1977. Pub. No. 15. Bibliografía bibliog.pmd 117 13/07/2005, 12:35 117 West, N.E. 1988. Intermountain deserts, shrub steppes and woodlands. En: M.G. Barbour y W.D. Billings (eds.). North American terrestrial vegetation. Cambridge University Press, Cambridge. Pp. 209-230. Wiggins, L.I. 1980. Flora of Baja California. Stanford University Press, Stanford, California. Woodward, I. F. 1987. Climate and plant distribution. Serie Cambridge Studies in Ecology. Cambridge University Press.Cambridge, Great Britain. 174 pp. (http:/ referente.allrefer.com/wildlife-plants-animals/plants/tree/). Zañoni, A.T. y R.P. Adams. 1973. Distribution and synonymy of Juniperus californica (Cupressaceae) in Baja California, México. Bulletin of Torrey Botanical. Club 100(6): 364-367. Yeaton, R.I. 1983. The succesional replacement of ponderosa pine by sugar pine in the Sierra Nevada. Bulletin of Torrey Botanical Club 110(3): 292-297. 118 El bosque de coníferas bibliog.pmd 118 13/07/2005, 12:35 Anexo I. Catálogo florístico principal de la flora vascular en el bosque de coníferas de la Sierra de San Pedro Mártir, Baja California División Lycopodiophyta Woodsia oregano D. Eaton Woodsia plummerae Lemmon Equisetaceae Woodwardia fimbriata Smith Equisetum hyemale L. ssp. affine (Engelm.) Calder & R.H. Taylor División Pinophyta Equisetum laevigatum A. Braun ORDEN GNETALES Selaginellaceae Ephedraceae Selaginella eremophylla Maxon Ephedra californica S. Watson División Polypodiophyta ORDEN PINALES Polypodiaceae Cupressaceae Cheilanthes lindheimeri (J. Smith) Hook Calocedrus decurrens (Torrey) Florin Cheilanthes wootonii Maxon Cupressus montana Wiggins Pellaea mucronata D. Eaton Juniperus californica Carriere Pellaea ternifolia (Cav.) Link ssp. wrightiana (Hook) Tryon Pinaceae Pentagramma triangularis (Kaulf ) G. Abies concolor (Gordon & Glerd) Lindley Yatskievych, M.D. Windham & E. Wollen. Pinus contorta Loudon Polypodium hesperium Maxon Pinus jeffreyi Grev & Balf. Pteridium aquilinum (L.) Kuhn var. Pinus lambertiana Douglas pubescens L. Underw. Pinus monophylla Torrey & Fremont Thelypteris puberula (Baker) C. Morton Pinus quadrifolia Parl. var. sonorensis A. Reed Smith Catálogo florísitico 119 Division Magnoliophyta Asteraceae CLASE MAGNOLIOPSIDA Achillea millefolium L. Agoseris heterophylla (Nutt.) E. Greene Amaranthaceae Antennaria parvifolia Nutt. Amaranthus blitoides S. Watson Artemisia douglasiana Besser Artemisia dracunculus L. Anacardiaceae Rhus kearneyi var. kearneyi Barkl. Artemisia ludoviciana Nutt ssp. albula (Woot.) Keck Rhus ovata S. Watson Artemisia tridentata Nutt. Rhus trilobata Torrey & A. Gray Aster occidentalis (Nutt.) Torr & A. Gray Rhus trilobata Nutt ex Torr & Gray var. simplicifolia (Greene) Barkley var. delectabilis (H.M. Hall) Ferris Bahia dissecta (A. Gray) Britton Baileya pleniradiata A. Gray Apiaceae Bidens heterosperma A. Gray Apium graveolens L. Blepharoneuron tricholepis (Torr.) Nash Berula erecta (Hudson) Cov. Brickellia grandiflora (Hook.) Nash Caucalis microcarpa Hook. & Arn. Brickelia sessile B. L. Robinson Cicuta maculata var. Maculata L. Centaurea melitensis L. Heracleum maximum Bartr. Cirsium californicum A. Gray Hydrocotyle ranunculoides L. F. Cirsium foliosum (Hook.) DC. Lomatium dasycarpus (Torrey & A. Gray) Cirsium scariosum Nutt. J. Coulter & Rose Lomatium lucidum (Torrey & A. Gray) Jepson Cirsium tioganum (Congdom) Petr. Cirsium trachylomum S. F. Blake Cirsium vulgare (Savi) Ten. Sanicula crassicaulis DC. Coniza canadensis (L.) Cronq. Sphenosciadum capitellatum A. Gray Conyza schiedeana (Less.) Cronquist Chaenactis parishii A. Gray Apocynaceae Apocynum cannabinum var. glaberri mum L. Apocynum sibiricum Jacq. Chrysothamnus nauseosus (Pallas) Britton ssp. consimilis (E. Greene) H.M. Hall & Clements Ericameria arborescens (A. Gray) E. Greene ssp. peninsularis Moran Asclepiaceae Ericameria ericoides (Less.) Jepson Asclepias fascicularis Decne. Ericameria linearifolia (DC.) Urb. & J. Wussow Ericameria martirensis Wiggins 120 El bosque de coníferas Ericameria parishii (Greene) H. M. Mall. var. peninsularis (Moran) G. L. Nesom Hemizonia martirensis Keck Heterotheca martirensis Moran Erigeron canadensis Brot. Hieracium albiflorum Hook Erigeron discoideus Kellog. Hieracium fendleri Sch. Bip. Erigeron divergens Torrey & A. Gray var. Hofmeisteria pluriseta A. Gray divergens Erigeron divergens Torrey & A. Gray var. cinereum A. Gray Erigeron foliosus Nutt. var. stenophyllus (Nutts) & A. Gray Erigeron foliosus Nutt. var. foliosus Nutt. Hulsea mexicana Rydb. Hymenopappus filifolius (Hook) var. lugens (E. Greene) Jepson Hymenothrix wrightii A. Gray Hypericum anagalloides Cham. & Schlecht. Erigeron formosissimus Greene Hypericum formossum var. scouleri Takht. Erigeron speciosus (Lindl.) DC. var. Isocoma menziesii (Hook. & Arn.) G. macranthus Cronquist Eriophyllum wallacei (A. Gray) A. Gray Eupatorium herbaceum Greene Filago californica Nutt. Geraea viscida (A. Gray) S.F. Blake Nesom var. menziesii Layia platyglossa var. campestris (Fischer y C. Meyer) A. Gray Machaeranthera juncea (E. Greene) Shinn Gnaphalium bicolor Bioletti Machaeranthera pinnatifida (Hook) Gnaphalium chilense Spreng. Shinn. var. gooddingii (Nelson) B. Turner Gnaphalium microcephalum Nutt. & R. Hartman Gnaphalium palustre Nutt. Machaeranthera tephrodes Greene Gutierrezia bracteata Abrams Machaeranthera wigginsii (S.F. Blake) R.L. Gutierrezia californica (DC.) Torrey & A. Gray Gutierrezia sarothrae (Pursh) Britton & Rusby Hartm Malacothrix clevelandii A. Gray Malacothrix glabrata A. Gray Matricaria matricarioides (Less.) Porter Haplopappus pulvinatus Moran Pectis papposa Harvey & A. Gray Haplopappus spinulosus Pursh) DC. ssp. Rafinesquia californica Nutt. gooddingii (A. Nels) Hall Senecio moranii T.M. Barcleq. Hazardia squarrosa (Hook & Arn.) E. Senecio spartioides Torrey & A. Gray Greene var. grindelioides (DC.) W. Clark Solidago californica Nutt. Helianthus californicus DC. Solidago sphatulata DC. var. Helenium puberulum DC. Hemizonia fasciculata (DC.) Torrey & A. Gray neomexicana (Gray) Cronq Solidago neomexicana Wooton & Standley Catálogo florísitico 121 Sonchus asper (L.) Hill Sphaeromeria martirensis (Wiggins) Holmgren, Schultz & Lowery Stephanomeria exigua Nutt. var. deanei Plagiobothrys californicus (A. Gray) Greene Plagiobothrys collinus var. fulvescens (I. M. Johonston) Higgins (J.F. Macbr.) Gottlieb Stephanomeria monocephala Moran Brassicaceae Stephanomeria pauciflora (Nutt.) Nelson Arabis perennans S. Watson Stephanomeria tenuifolia (Torrey) H.M. Barbarea orthoceras Leden. Hall Stephanomeria virgata Benth var. pleurocarpus (E. Greene) Gottlieb. Taraxacum laevigatum D.C. Taraxacum officinale Wigg. Tessaria sericea (Nutt.) Shinners Uropappus lindleyi (DC.) Nutt Verbesina palmeri S. Wats Xanthisma wigginsii (S.F. Blake) D. R. Moran & R. L. Hartm. Xylothamia diffusa (Benth.) G. L. Nesom Descurainea obtusa (E. Greene) O. Schulz var. obtusa Descurainea pinnata (Walter) Britton var. halictorum (Cockerell) Detl. Descurainea pinnata (Walter) Britton var. glabra (Wooton & Standley) Detl. Descurainea richardsonii var. viscosa (Sweet) Schulz Diplotaxis muralis (L.) DC. Draba corrugata S. Watson var. demareei (Wigg) C.L. Hitchc. Lesquerella peninsularis Wigg. Berberidaceae Berberis higginsae Munz Lepidium lasiocarpum Nutt. ex Torr. & A. Gray var. lasiocarpum Lepidium lasiocarpum Nutt. ex Torr. & A. Boraginaceae Cryptantha clevelandii E. Greene Cryptantha micrantha (Torrey) I.M. Johnston var. lepida (A. Gray) I.M.Johnston Gray var. latifolium C.L. Hitchc. Lepidium virginicum L. var. robinsonii (Thell.) C. Hitchc. Lepidium virginicum L. var. pubescens (E. Greene) Thell Cryptantha muricata (Hook & Arn.) Pennellia micrantha Nieuwl. Nelson & J.F. Macbr. Rorippa curvisiliqua (Hook.) Bessey ex Cryptantha muricata (Hook. & Arn.) A. Nelson & J.F. Macbr. var. denticulata (Greene) I.M. Johnst. Hackelia pinetorum (Greene ex A. Gray) I.M. Johnst. var. pinetorum Pectocaria recurvata I.M. Johnston 122 El bosque de coníferas Britton var. orientalis Stuckey Rorippa nasturtium-aquaticum (L.) Hayek Rorippa tenerrima E. Greene Streptanthus campestris S. Watson Thelypodium wrightii A. Gray Hutchinsia procumbens (L.) Desv. Silene verecunda S. Watson ssp. platyota (S. Watson) C. Hitchc. & Maquire Cactaceae Ceratophyllaceae Echinocereus engelmannii (Engelm.) Ceratophyllum demersum L. Lemaire Echinocereus pacificus Britton & Rose Chenopodiaceae Opuntia californica ( Torr. & A. Gray) Chenopodium berlandieri Moq. Coville var. parkeri (J.M. Coult.) Pinkava Chenopodium fremontii S. Watson Opuntia ganderi (C.B. Wolf ) J. Rebman & Chenopodium incanum (S. Watson) A.A. D. J. Pinkava var. ganderi Heller ChenopChenopodium graveolens Willd. Campanulaceae var. neomexicanum Aellen Heterocodon rariflorum Nutt. Chenopodium leptophyllum Moq. Lobelia cardinalis L. ssp. graminea (Lam.) MC Vaugh Commelinaceae Lobelia dunnii E. Greene var. dunnii Tripogandra angustifolia (B.L. Robinson) Woodson Caprifoliaceae Lonicera subspicata Hook. & Arn. var. Convolvulaceae johnstonii D. D. Keck Convolvulus aridus Sambucus mexicana C. Presl ex DC. tenuifolius Abrams Greene ssp. Symphoricarpus longiflorus A. Gray Symphoricarpus rotundifolius A. Gray var. Crassulaceae parishii (Rydb.) Dempster Dudleya pauciflora Rose Sedum niveum Davidson Caryophyllaceae Arenaria confusa Rydb. Cuscutaceae Drymaria leptophylla (Cham. & Schl.) Cuscuta californica Hook & Arn. Fenzl ex Rohrb. var. leptophylla Cuscuta ceanothi Behr. Drymaria tenella A. Gray Sagina decumbens (Elliott) Torrey & A Datiscaceae Gray Datisca glomerata (C. Presl.) Baillon Sagina saginoides (L.) Karsten Silene laciniata Cav. ssp. brandegeei Ericaceae Hitch. & R. Moran Arctostaphylos glauca Lindl. Silene laciniata Cav. var. major C. Hitchc. Arctostaphylos patula Greene ssp. Catálogo florísitico 123 platyphylla (A. Gray) P. V. wells. Lotus scoparius (Nutt. ex Torr. & A. Gray) Arctostaphylos peninsularis P. V. Wells. Ottley var. brevialatus (Ottley) Munz Arctostaphylos pringlei Parry ssp. Lotus strigosus (Nutt.)E. Greene var. drupacea hirtellus (E. Greene) Ottley Arctostaphylos pungens Kunth. Lotus strigosus (Nutt.) E. Greene var. strigosus Euphorbiaceae Lotus utahensis Ottley Acalypha californica Benth Lupinus andersonnii S. Watson var. Bernardia incana Morton sublinearis Chaenactis parishii A. Gray Lupinus concinnus (S. Watson) C.P. Smith Chamaesyce melanadenia (Torr.) Millsp. Lupinus excubitus M.E. Jones var. Croton californicus Muell austromontanus (A.A. Heller) C.P. Smith Euphorbia micromera Boiss Lupinus hyacinthinus E. Greene Euphorbia palmeri Engelm. Lupinus latifolius J. Agardh var. wigginsii Euphorbia serpyllifolia Pers. Lupinus truncatus Nutt. ex Hook. & Arn. var. truncatus Fabaceae Marina orcuttii (S. Watson) Barneby Amorpha apiculata Wiggins Medicago hispida Gaertn. Amorpha californica Nutt. Medicago polymorpha L. Astragalus circumdatus Greene Melilotus indicus (L.) All Astragalus douglasii ( Torrey & Gray) Mimosa biuncifera Benth. A.Gray var. parishii (A.Gray) M.E. Jones Psoralea rigida Parish Astragalus gruinus Barneby Trifolium microcephalum Pursh Astragalus palmeri A. Gray var. palmeri Trifolium variegatum Nutt. Astragalus prorifer M.E. Jones Trifolium wigginsii J.M. Gillett Hoita orbicularis (Lindley) Rydb Trifolium wormskioldii Lehm Lathyrus splendens Kellogg Vicia americana Willd var. linearis S. Lotus argyraeus (Greene) Greene Watson Lotus crassifolius (Benth.) Greene var. crassifolius Fagaceae Lotus grandiflorus (Benth) E. Greene var. Quercus agrifolia Nee var. oxidenia anthyloide (Torrey) J. Howell Lotus humistratus E. Greene Quercus chrysolepis Liebm. Lotus nevadensis (S. Watson) E. Greene Quercus dumosa Nutt. Lotus oblongifolius (Benth) E. Greene Quercus peninsularis Trel. Lotus rigidus (Benth) E. Greene Quercus turbinella E. Greene 124 El bosque de coníferas Quercus wislizenii A. DC. var. frutescens vón) Choisy Engelm Quercus wislizenii A. DC. var. wislizenii Lamiaceae Dracocephalum parviflorum Nutt. Garryaceae Hedeoma martirense Moran Garrya grisea Wiggins Marrubium vulgare L. Mentha arvensis L. Gentianaceae Mentha spicata L. Centaurium exaltatum (Griseb.) Piper Monardella lanceolata A.Gray Frasera parryi Torr. Monardella linoides A. Gray var. viminia Gentianella amarella (L.) Boerner spp. (E. Greene) Abrams acuta (Michaux) J.M. Gillett Monardella macrantha A. Gray Swertia parryi (Torrey) Kuntze Poliomintha conjunctrix Epling. & Wigg. Prunella vulgaris L. var. lanceolata Geraniaceae (Barton) Fern. Erodium cicutarium (L.) L’Her. Salvia pachyphylla Munz Geranium atropurpureum A. Heller Stachys ajugoides Benth. var. rigida Geranium gracile Ledeb. ex Nordm. Jepson & Hoover Grossulariaceae quercetorum (A. Heller) Epling Ribes quercetorum E. Greene Teucrium glandulosum Kellogg. Stachys rigida Nutt. ex Benth. ssp. Trichostemma lancelatum Benth Hydrophyllaceae Trichostemma parishii Vasey Eriodictyon angustifolium Nutt. Nama dichotomum (Ruiz López & Pavón) Lemnaceae Choisy Lemna minor L. Phacelia affinis A. Gray Lemna trisulca L. Phacelia brachyloba (Benth) A. Gray Phacelia cryptantha E. Greene Lennoaceae Phacelia distans Benth Pholisma arenarium Hook. Phacelia imbricata E. Greene var. patula (Brand) Heckard Linaceae Phacelia mutabilis E. Greene Linum australe A. Heller Phacelia parryi Torrey Linum lewisii Pursh Phacelia pedicellata A. Gray Linum neomexicanum Greene Phacelia sufrutescens (Ruiz López & Pa- Catálogo florísitico 125 Lythraceae Dendromecon rigida Benth var. rigida Lythrum californicum Torr. & A. Gray Malvaceae Plantaginaceae Malacothamnus fasciculatus (Torrey & A. Plantago erecta E. Morris Gray) E. Greene Plantago linearis Kunth var. mexicana Sidalcea malvaeflora Benth ssp. (Link) Pilg. sparsifolia C. Hitchc Plantago major L. Onagraceae Polemoniaceae Cammisonia californica ( Torrey & A. Eriastrum diffusum (A. Gray) H. Mason Gray) Raven Eriastrum densifolium (Benth) H. Mason Camissonia graciliflora (Hook. & Arn.) P.H. Eriastrum densifolium (Benth) H. Mason Raven var. elongatum Camissonia hiertella (E. Greene) Raven Gilia mexicana A.D. Grant & V.E. Grant Camissonia ignota (Jepson) Raven Ipomopsis efusa (A. Gray) Moran Camissonia pallida (Abrams) Raven var. Ipomopsis tenuifolia (A. Gray) V. Grant pallida Linanthus floribundus (A. Gray) Milliken Camissonia strigulosa (Fischer& C. Linanthus floribundus (A. Gray) Milliken Meyer) Raven ssp. Glaber R. Patterson Epilobium adenocaulon Hausskn. var. Linanthus melingii (Wiggins) V.E. Grant parishii (Trel.) Munz Linanthus orcuttii (Parry & A. Gray) Epilobium glaberrimum Barbey in Jepson Brewer & S. Watson ssp. glaberrimum Linanthus pygmaeus (Brand) J. Howell Epilobium canum (E. Greene) Raven Linanthus pygmaeus (Brand) J.T. Howell Epilobium ciliatum Raf. ssp. ciliatum ssp. continentalis Raven Gayophitum diffusum Torrey & A. Gray Navarretia atractyloides (Benth) Hook. & Oenothera californica (S. Watson) S. Arn. Watson Phlox austromontana Cov. Oenothera elata var. hirsutissima Kunth Polygonaceae Orobanchaceae Chorizanthe fimbriata Nutt var. fimbriata Orobanche parishii (Jeps.) Heckard var. Eriogonum elongatum parishii elongatum Benth var. Eriogonum fasciculatum Benth var. Papaveraceae polifolium (A. DC.) Torrey Argemone subintegrifolia G.B. Ownbey Eriogonum fasciculatum Benth var. 126 El bosque de coníferas foliosum Zannichellia palustris L. Eriogonum foliosum S. Watson var. hastatum Pyrolaceae Eriogonum gracile Benth Pterospra andrómeda Nutt. Eriogonum hastatum Wigg. Eriogonum nudum Benth Sarcodes sanguinea Torr. var. pauciflorum S. Watson Ranunculaceae Eriogonum parishii S. Watson Aquilegia formosa Fisher var. hypolasia (E. Eriogonum thurberi Torrey Eriogonum wrightii Greene) Munz Benth var. Clematis pauciflora Nutt membranaceum Jepson Clematis ligusticifolia Nutt Eriogonum wrightii Benth var. oresbium Ranunculus aquatilus L. var. capillaceus Eriogonum (Thuill.) DC. wrigthii Benth var. subscaposum S. Watson Ranunculus cymbalaria Pursh var. Polygonum amphibium L. Coleman var. saximontanus Fern. stipulaceum Ranunculus uncinatus D.Don Polygonum arenastrum Boreau Thalictrum fendleri A. Gray var. fendleri Polygonum douglasii E. Greene var. johnstonii (Munz) J. Hickman Rhamnaceae Pterostegia drymarioides Fisher y C. Ceanothus cordulatus Kellog Meyer Ceanothus cuneatus (Hook) Nutt Rumex californicus Rech. F. Ceanothus greggii A. Gray var. perplexans Rumex crispus L. (Treel) Jepson Rumex salicifolius Weinm var. salicifolius Ceanothus leucodermis E. Greene Rumex Ceanothus spinosus Torrey & A. Gray salicifolius Weinm var. denticulatus Torr. Ceanothus verrucosus Nutt Rhamnus californica Eschsch ssp. Portulacaceae californica Calyptridium monandrum Nutt Rhamnus californica Eschsch ssp. ursina Calyptridium umbellatum (Torrey) E. (Greene) C.B. Wolf Greene Rhamnus ilicifolia Kellogg Portulaca oleracea L. Rhamnus tomentella Benth ssp. tomentella Potamogenaceae Potamogeton pectinatus L. Rosaceae Potamogeton pusillus L. Adenostoma fasciculatum Hook & Arn Catálogo florísitico 127 Adenostoma sparsifolium Torrey Galium wrightii A. Gray Amellanchier utahensis Koehne Cercocarpus betuloides Torrey & A. Gray Salicaceae Cercocarpus ledifolius Nutt Populus tremuloides Michaux. Fragaria californica Cham. & Schltdl. Salix lasiolepis Benth. Heteromeles arbutifolia (Lindley) Roemer Holodiscus dumosus (S. Watson) A. Heller Saururaceae Holodiscus microphyllus Rydb var. Anemopsis californica (Nutt) Hook & Arn. microphyllum Holodiscus microphyllus Rydb var. sericeus F.A. Ley Saxifragaceae Horkelia bolanderi A. Gray Heuchera leptomeria Greene var. Ivesia argyrocoma (Rydb.) Rydb. peninsularis Potentilla glandulosa Lindley ssp. reflexa Heuchera rubescens Torrey var. versicolor (E. Greene) Keck (E. Greene) M. G. Stewart Potentilla gracilis Douglas ex Hook ssp. Philadelphus microphyllus A. Gray var. nuttallii (Lehm.) D.D. Keck stramineus (Rydb.) C.L. Hitchc. Potentilla rimicola (Munz & J. M Saxifraga arguta D. Don Johnston) B. Ertter Saxifraga eriophora S. Watson Potentilla wheeleri S. Watson Prunus emarginata (Hook) Walp. Scrophulariaceae Prunus fasciculata (Torrey) A. Gray Antirrhinum coulterianum Benth var. Rosa californica Cham. & Schldl coulterianum (A. Gray) Munz Castilleja applegatei Fern ssp. martinii Rubiaceae (Abrams) Chaung & Heckard Galium andrewsii A. Gray Castilleja minor (A. Gray) A. Gray ssp. Galium angustifolium Nutt. ex A. Gray var. spiralis (Jepson) Chuang y Heckard angustifolium Castilleja stenantha A. Gray Galium angustifolium Nutt. ex A. Gray var. Cordylanthus involutus Wiggins diffusum Hilend & J.T. Howell Cordylanthus rigidus (Benth) Jepson ssp. Galium boreale L. var. septentrionale involutus ( Wiggins) T.I. Chuang & (Roem. & J. A. Schultes) H. Hara Heckard Galium diabloense Dempster Cordylanthus nevinii A. Gray Galium martirense Dempster & Stebbins Keckiella antirrhinoides (Benth.) Straw Galium nuttallii A. Gray ssp. nuttallii var. antirrhinoides Galium wigginsii Dempster Keckiella ternata (Torr. ex A. Gray) Straw 128 El bosque de coníferas var. ternata Viscaceae Limosella acaulis Sesse & Mociño Arceuthobium americanum Engelm. Mimulus brevipes Benth Arceuthobium californicum Hawksw. & Mimulus cardinalis Benth Wiens Mimulus floribundus Lindley Arceuthobium campylopodium Engelm. Mimulus guttatus DC. Phoradendron densum Trel. Mimulus nasutus Greene Phoradendron libocedri (Engelm.) Mimulus pilosus (Benth) S. Watson Howell Mimulus purpureus var. pauxillus A. L. Phoradendron macrophyllum (Engelm.) Grant Cockerell. Penstemon californicus (Munz & I. M. Phoradendron pauciflorum Torr. Johnston) Heck Phoradendron villosum Nutt. ssp. Penstemon centranthifolius (Benth) villosum Benth Penstemon clevelandii A. Gray var. CLASE LILIOPSIDA clevelandii Penstemon spectabilis Thurber ex Gray Agavaceae Penstemon thurberi Torrey Nolina palmeri S. Watson var. palmeri Veronica Americana (Raf.) Schwein Yucca schidigera K. E. Ortgies Veronica peregrina L. var. xalapensis Yucca whipplei Torrey (Kunth) Pennell Veronica serpillifolia L. var. humifusa Amaryllidaceae (Dickson) Syme. Muilla maritima (Torr.) S. Watson Solanaceae Cyperaceae Nicotiana attenuata Torrey Carex agrostoides Mack Solanum sarachoides Sendtner Carex athrostachya Olney. Solanum xantii A. Gray Carex douglasii Boott Carex globosa Bott Sterculiaceae Carex lanuginosa Michx. Fremotodendron californicum (Torrey) Cov. Carex praegracilis W. Boott Carex rostrata Stokes Urticaceae Carex senta Boott Urtica dioica L. var. holosericea (Nutt) Carex subfusca W. Boott Thorne Cyperus niger Ruiz López & Pavón var. capitatus Catálogo florísitico 129 Cyperus squarrosus L. Orchidaceae Eleocharis acicularis (L.) Roemer & Epipactis gigantean Hook. Schultes Platanthera leucostachya Lindsay Eleocharis bella Piper Platanthera sparsiflora (S. Watson) Schltr. Eleocharis macrostachya Britton Eleocharis montevidensis Kunth Poaceae Eleocharis parishii Britton Agropyron trachycaulum (Link) Malte Scirpus americanus Pers. Agropyron trachycaulum (Link) Malte var. unilaterale (Cassidy) Malte Iridaceae Agrostis exarata Trin. Sisyrinchium bellum S. Watson Agrostis scabra Willd. Agrostis semiverticillata (Forssk.) Christ. Juncaceae Agrostis stolonifera L. var. palustris Juncus articulatus L. (Huds.) Farw. Juncus balticus Willd. Agrostis stolonifera L. Juncus bryoides F. J. Herm. Alopecurus aequalis Sobol. Juncus bufonius L. Andropogon glomeratus (Walter) Britton, Juncus dubius Engelm. Sterns & Pogg. Juncus effusus L. var. pacificus Fern. Aristida schiediana Trin. & Rupr. Juncus kelloggii Engelm. Blepharoneuron tricholepis (Torr.) Nash. Juncus macrophyllus Cov. Bouteloua curtipendula (Michaux) Torrey Juncus mexicanus Willd. Bromus carinatus Hook. & Arn. Juncus oximeris Engelm. Bromus ciliatus L. Juncus rugulosus Engelm. Bromus diandrus Roth. Juncus saximontanus Nelson Bromus madritensis L. ssp. rubens (L.) Juncus tiehmii B. Ertter Husnot Juncus xiphioides E. Meyer Bromus tectorum L. Luzula comosa E. Meyer Deschampsia caespitosa (L.) Beauv. ssp. caespitosa Liliaceae Deschampsia elongate (Hook.) Benth. Allium haematochiton S. Watson Dichanthelium oligosanthes (Schult.) Calochortus concolor (Baker) Purdy Gould var. scribnerianum (Nash) Gold Elymus elymoides (Raf.) Swezey ssp. Calochortus splendens Benth. Calochortus weedii Wood. peninsularis Ownbey var. elymoides Elymus longifolius (J. G. Sm.) Gould Elymus multisetus (J. G. Sm.) Davy 130 El bosque de coníferas Eragrostis intermedia Hitchc. Muhlenbergia wolfii (Vasey) Rydb. Eragrostis pectinacea (Michaux.) Nees Muhlenbergia wrightii Vasey Festuca ovina L. Paspalum paspalodes (Michx.) Scibn. Hordeum brachyanthemum Nevski Poa fendleriana (Steud.) Vasey Hordeum glaucum Steud. Poa orcuttiana Vasey Hordeum jubatum L. Poa pratensis L. ssp. pratensis Koeleria macrantha (Ledeb.) J. A. Polypogon monspelliensis (L.) Desf. Schultes Polypogon semiverticellata (Forssk.) Koeleria pyramidata (Lam.) Beauv. Christ. Leptochloa dubia (H.B.K.) Ness Sphenopholis obtusata (Michx) Scribn. Lycurus phleoides Kunth var. phleoides Sporobulus criptandrus (Torr.) Gray Muhlenbergia arsenei A. Hitchc. Stipa coronata Thurb. Muhlenbergia asperifolia (Ness & Meyen) Stipa lepida Hitchc. Var. lepida L. Parodi Stipa pringlei (Beal) Scibn. Muhlenbergia emersleyi Vasey Stipa speciosa Trin. & Rupr. Muhlenbergia filiformis (Thurbey) Rydb. Vulpia myurus (L.) C. C. Gmelin var. hirsu- Muhlenbergia fragilis Swallen ta Hack. Muhlenbergia minutissima (Steud) Vulpia octoflora (Walt.) Rydb var. glauca Swallen (Nutt.) Fern. Muhlenbergia pauciflora Buckley Vulpia octoflora (Walt.) Rydb var. hirtella Muhlenbergia repens (Presl.) Hitchc. (Piper) Hern. Muhlenbergia richardsonniis (Trin.) Rydb. Vulpia octoflora ( Walt.) Rydb.var. Muhlenbergia rigens (Benth.) Hitchc. octoflora Catálogo florísitico 131 132 El bosque de coníferas Anexo 2. Catálogo fotográfico Coníferas Abies concolor Conos de A. concolor Catálogo fotográfico 133 Calocedros decurrens Pinus jeffreyi 134 El bosque de coníferas Corteza de P. jeffreyi (“viejo”) Corteza de P. jeffreyi (“joven”) Hojas y conos de P. jeffreyi Conos de P. jeffreyi Catálogo fotográfico 135 Pinus lambertiana Corteza y conos de P. lambertiana Pinus contorta ssp. murrayana Corteza de P. contorta ssp.murrayana 136 El bosque de coníferas Cono de P. contorta ssp.murrayana Cupressus montana Catálogo fotográfico 137 Pinus coulteri Pinus monophylla 138 El bosque de coníferas Conos de Pinus monophylla Hojas de Pinus monophylla Pinus quadrifolia Cono de P. quadrifolia Catálogo fotográfico 139 Juniperus californica 140 El bosque de coníferas Árboles (noconíferas) Populus tremuloides (alamillo) Hojas y tronco de P. tremuloides Catálogo fotográfico 141 Salix lasiolepis (sauce) Quercus chrysolepis 142 El bosque de coníferas Coníferas Adenostoma sparsifolium (chamizo rojo) Catálogo fotográfico 143 Symphoricarpus mollis Nutt. Eriogonum wrigthij var. subscaposum 144 El bosque de coníferas Arctostaphylos patula (manzanita) Arctostaphylos peninsulares (manzanita) Hojas de A. patula Ceanothus greggii var. perplexans Ceanothus leucodermis Catálogo fotográfico 145 Garrya grisea Rhamnsu californica ssp. californica 146 El bosque de coníferas Ericameria parishii var. peninsulares Nolina palmeri var. palmeri Catálogo fotográfico 147 Herbáceas Achillea millefolium var. californica Penstemon centranthifolius 148 El bosque de coníferas Monardella macrantha Pteridium aquilinum var. lanuginosum (helecho) Catálogo fotográfico 149 Astragalus douglasii var. glaberrimus Hulsea mexicanum 150 El bosque de coníferas Erigeron divergens var. cinereum Euphorbia palmeri Catálogo fotográfico 151 Trifolium wigginsii Salvia pachyphylla 152 El bosque de coníferas Potentilla wheeleri Silene laciniata ssp. brandegeei Catálogo fotográfico 153 Linanthus melingii Lotus scoparius var. scoparius 154 El bosque de coníferas Lupinus hyacinthinus Catálogo fotográfico 155 Lupinus excubitus ssp. austromontanus Lupinus truncatus 156 El bosque de coníferas Phoradendron bolleanum ssp. pauciflorum Planta parásita de Abies concolor Catálogo fotográfico 157 Sarcodes sanguinea Planta saprofita en raíces de coníferas 158 El bosque de coníferas Catálogo fotográfico 159 El bosque de coníferas de la Sierra de San Pedro Mártir, Baja California, México de José Delgadillo Rodríguez se terminó de imprimir durante el mes de diciembre de 2004 en los talleres gráficos de la empresa Master Print en la Ciudad de México. Se tiraron 700 ejemplares 160 El bosque de coníferas